
Taxonomy

Our understanding of species concepts goes to the very heart of the way Nature is organised. It is endlessly complex and is probably a little different for each and every species. Both practical and theoretical issues arise from it that have broad relevance to the taxonomy, biogeography, systematics and evolutionary concepts of most marine organisms, not just corals.

Theoretical background
Species have long been considered to be the ‘building blocks’ of Nature, blocks that have a time and place of origin, which can evolve, and which maintain themselves as discrete entities by being reproductively isolated. This is a logical concept, but one which breaks down, for most corals, when confronted with the realities of geographic variation. The alternative, where most species have none of these attributes, is initially less intuitive but ultimately is highly explanatory of what can actually be observed in the real world.
Concepts of what species are have long been debated, a debate driven as much by misinterpretation as information. The debate, however, has mostly been ignored by taxonomists (who often believe they ‘know’ what their species are for one reason or another) as well as most other biologists (who often consider it ‘armchair philosophy’ and thus largely irrelevant to the needs of reality). Be that as it may, species have variously been considered to be (a) self-defining natural units, (b) human-defined units composed of assemblages united by common descent, or (c) genetically-defined units resulting from Darwinian natural selection and/or genetic (reproductive) isolation. The last involves a group of concepts, sometimes called the ‘neo-Darwinian synthesis’ or the ‘biological species concept’ (depending on a wealth of detail) which embodies the notions of the building blocks referred to above.
Syngameons
The concept of the syngameon, first recognised by botanists and introduced to the marine world through corals, is important for the understanding of the evolutionary mechanisms of corals and the geographic patterns they form. A syngameon, by definition, is a reproductively isolated unit. In concept, so are neo-Darwinian species. In reality, syngameons are nothing like any of these concepts of species as they incorporate geographic variation and the spectrum of genetic links geographic variation creates among different species.
A syngameon may be a single species, or it may be a cluster of different species which have variable genetic links (genetic flow through cross-fertilisation) with other members of the syngameon. Where a syngameon contains several species, a single component species may be distinct at a single geographic location but, because it intergrades with other species at other locations, it will become submerged in a mosaic of variation at other locations. The geographic range and morphological variation of the single ‘species’ will depend on taxonomic decisions. These decisions will be arbitrary if they impose divisions on natural continua rather than reflect natural units. The syngameon as a whole is not morphologically distinguishable unless its component species are determined genetically or experimentally (through cross-fertilisation trials) in every part of the species’ possible distribution range.
Variation in species
In considering what species are it is helpful to compare corals with plants. Like most plants, corals have morphological characteristics that vary according to the environment in which they occur, their geographic location, and genetic links among component populations.
Environmental and genetic variation
Most colonial corals show morphological variation within a single colony. Colonies of some species have variable growth forms which are at least partly genetically regulated independently of environment. However, most species exhibit variations that are clearly related to different micro-environments across the colony. Such differences include growing space, light availability, sedimentation and fish grazing. The morphological variations that result affect both growth form and corallite structure.

Echinopora mammiformis showing both flat plates and upright branches. Other colonies of the same species consist of plates only or branches only. Just as in plants and other colonial animals, single colonies of corals may have two or more distinct growth forms. Sometimes this variation appears to be controlled by genetic switches, turning different growth forms on or off. Philippines Photograph: Charlie Veron. Figure 1.
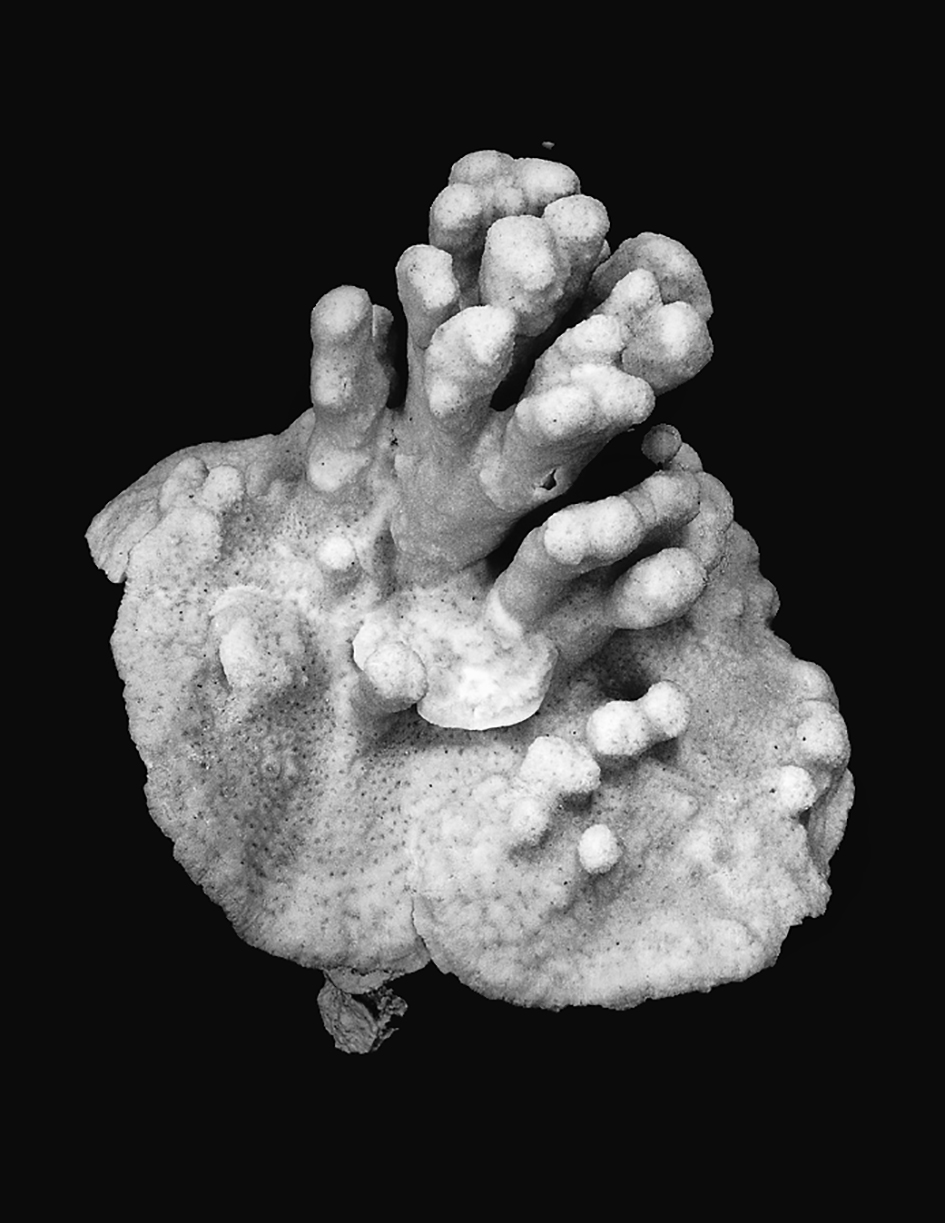
A small colony of Pavona maldivensis showing plate-like and columnar growth forms combined. These two growth forms are usually found in different colonies depending on the environment in which they occur. They were originally described as Siderastrea maldivensis Gardiner, 1905 and Pavona (Pseudocolumnastrea) policata Wells, 1954 and have been placed in different genera or subgenera by several taxonomists. Photograph: Veron archives. Figure 2.

Turbinaria mesenterina exposed to strong light. When exposed to strong light, T. mesenterina colonies such as this have highly convoluted growth forms. In progressively deeper or more turbid water where light levels become reduced, colonies become less convoluted until they are flat sheets. Papua New Guinea Photograph: Charlie Veron. Figure 3.
Different colonies (or individuals) of all coral species vary along environmental gradients. Divers can readily see this as they descend down a reef slope. This variation is also seen if a coral is transplanted from one habitat to another. It will usually change its appearance to reflect the conditions of the new habitat. Most plants do likewise; these responses are not under direct genetic control, although particular genotypes may be associated with particular morphologies.
The most important environmental factors controlling growth form are exposure to wave action, levels of illumination, sediment load and exposure to currents. The different morphologies that result must be accommodated by taxonomic descriptions and understood by persons identifying corals.
Geographic variation
There are two distinct categories of geographic variation in corals: those which are the outcome of environmental factors, and those which are genetic.
Geographic variation which is the outcome of environment is best seen in high latitude coral communities, where low temperature and non-reef habitats result in colonies which are distinct from their tropical reef-dwelling counterparts. Likewise, there are whole geographic regions where the water is almost always clear (such as the Red Sea or Bahamas) and other regions where the water almost always contains sediment from the land (such as the south-east Asian coast). Corals from these regions commonly have distinct points of morphological detail which are probably due to environmental differences.
Geographic variation which is genetically based affects almost all species in some way or other. Again like most plants, the appearance of a single species changes (gradually or abruptly) from one region to the next. These changes may not concern the individual who is interested in the species of a single country, but they will definitely concern anybody who tries to answer the question ‘what are species’? Of more practical importance, it will also concern anybody who tries to identify a species in one country from information about that species (including photographs) from another country. The taxonomic issues that arise are explained below, as these can also affect understanding of the species’ distribution range, crucial in conservation.
Genetic links among populations
It is common for a series of adjacent colonies of the same species growing in a uniform environment to display a wide range of colours and to have a variety of morphological differences. In such cases the presence or absence of morphological continuities among colonies, or populations, can be used to distinguish groups of species (commonly called ‘sibling species’) from a single variable species (commonly called ‘polymorphic species’). Although the differences seen are widespread, intermediate colonies are sometimes found. For this reason it is considered to be a single species.
Genetic bridges and barriers generate reticulate patterns in time and space, patterns which maintain the species’ genetic heterogeneity. The taxonomic issues that arise are of endless complexity. They inevitably lead to the conclusion that there are no fundamental differences between species and subspecies taxonomic levels.

Colonies of Lobophyllia hemprichii growing together commonly exhibit a wide range of colour and corallite shapes and structures. This is regarded as a single species because complete continua can be found among many colonies. Bali, Indonesia Photograph: Charlie Veron. Figure 4.

Montipora digitata: these two colonies on a reef flat of the Great Barrier Reef are considered the same species because colonies from other locations intergrade with them. Nevertheless, they represent populations with consistent differences. Great Barrier Reef, Australia Photograph: Ben Stobart. Figure 5.
Taxonomic issues
Before the advent of taxonomic studies on reefs, many coral taxonomists thought that a specimen with a different growth form implied a different species. Thus, thousands of supposedly ‘new species’ were described from specimens collected on expeditions and brought back to museums and universities for study. Scuba diving was largely responsible for changing this. It allowed taxonomists to observe variation in corals more or less as botanists had long observed variation in trees. It allowed differences between species that grow together to be studied in great detail and it also allowed environmental variation within the same species, such as occurs down a reef slope, to be observed directly.
Geographic variation is much less simple to study as knowledge of it must be accumulated from separate studies in different countries: it cannot be directly observed. The main taxonomic issues are: (a) species become progressively less recognisable as single units with increasing geographic range, (b) taxonomists are forced to make arbitrary decisions, (c) synonymies vary geographically. In each case, the more detailed a taxonomic study is, the greater the problem becomes.
Taxonomic certainty and geographic range
With corals as with most plants, most species do exist as more-or-less definable units in single geographic regions, such as the Red Sea, the Indonesian/Philippines archipelago, the Great Barrier Reef or the Caribbean. However, widespread species commonly show sufficient geographic variation that they could reasonably be divided into several separate ‘sibling species’ were it not for the fact that these smaller units form continua. Thus, for example, the majority of species of the Red Sea also occur in the central Indo-Pacific, but many are sufficiently different in the Red Sea that they would be considered to be distinct species if they were transplanted to the central Indo-Pacific.
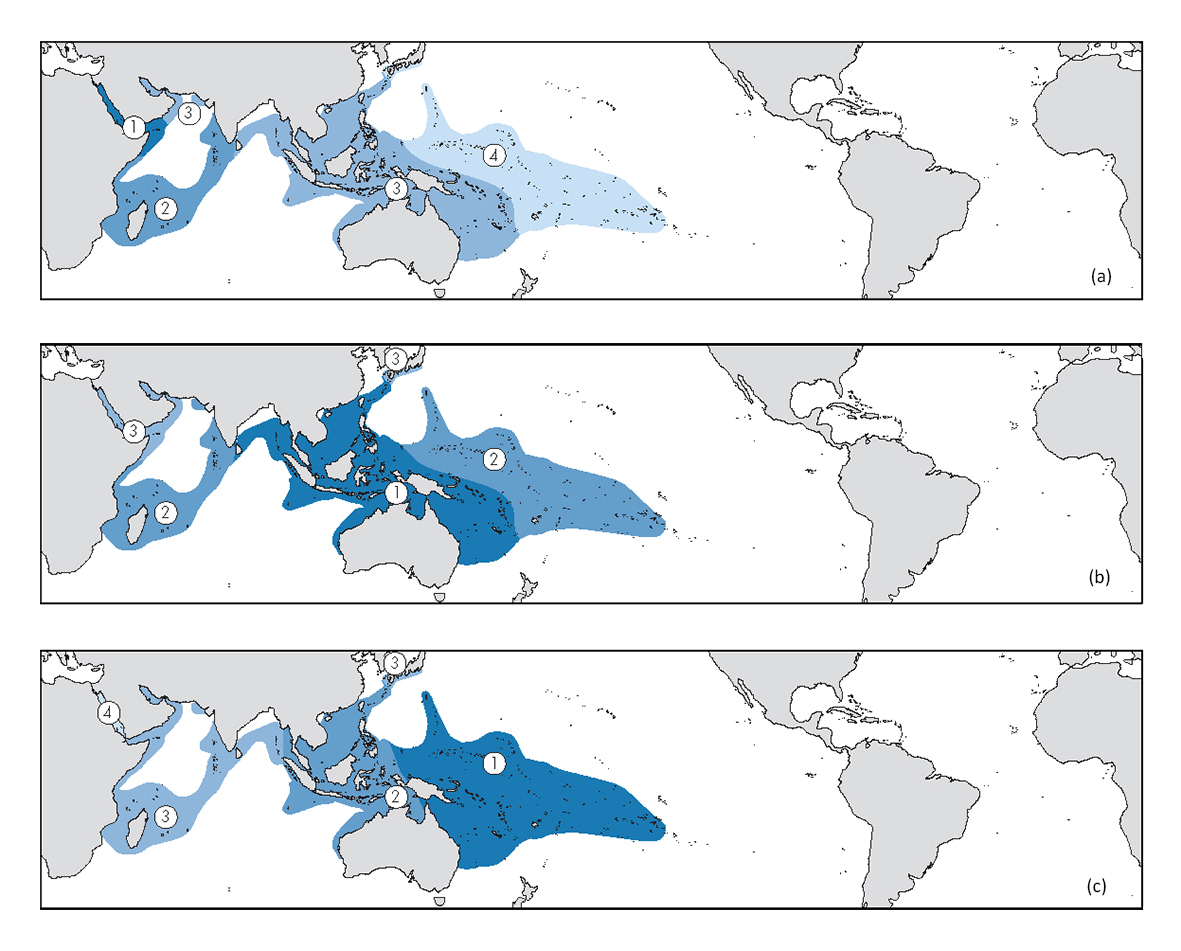
Taxonomists’ view of geographic variation. Corals, along with other tropical marine life, may have a west-to-east distribution range that spans much of the tropical Indo-west Pacific, a distance of half the circumference of the earth. Over such a vast distance, species gradually change. They may do so to the point where variation within a species becomes indistinguishable from variation between species. These maps show how the same species might appear to different taxonomists working primarily in (a) the Red Sea (top map), (b) the central Indo-Pacific (middle map) and (c) the south Pacific (bottom map). The taxonomist makes confident decisions in regions marked ‘1’, is uncertain in ‘2’, is very doubtful in ‘3’ and does not recognise the species in ‘4’. In general, the more widespread and detailed the taxonomic study, the less certain the taxonomist will be. From Veron (2000a) Corals of the World. Figure 6.
Arbitrary decisions
In theory, taxonomy should accommodate biogeographic patterns. In practice, doing this creates issues like those created when a flat map is projected onto a sphere. The bigger the area of the map, the greater the distortion that results. It is possible to modify the flat map using different types of projections, but this only changes the nature of the distortion.
In traditional taxonomy, geographic variation within a species is accommodated by creating divisions within the species, such as varieties, races or geographic subspecies. In reality, geographic variation repeatedly overrides the morphological boundaries of individual species. In other words, natural continua go beyond the taxonomic or morphological boundaries of individual species. This cannot be accommodated by creating divisions within species. The problem remains if the species unit is ‘split’ into smaller units or ‘lumped’ into larger units and it is not solvable by further or more detailed study. One example of this is the widespread species Micromussa amakusensis. Described originally from southern Japan, it has been recorded from the Indo-west Pacific including Arabia and the Red Sea (see its distribution factsheet). Consistent with reticulate patterns of evolution, the Arabian form shows some morphological and genetic differences from the Japanese population. Based on these differences the Arabian variant has been named Micromussa indiania by Arrigoni, Benzoni, Huang et al. (2016).
Ultimately, the only unit in Nature that is real is the syngameon, described above.
The reasons why syngameons are not used in operational taxonomy are: (a) they can only be determined with any degree of certainty through exhaustive cross fertilisation studies in all geographic regions where their component species occur; (b) they are not likely to have distinguishing morphological characteristics; and (c) they would include so many morphological species that they would need to be re-divided into sub-units of some kind in order to be useful.
These issues will always force taxonomists to make an arbitrary decision as to what a particular species is. Some groups of species may be distinctive in some geographic regions and not in others. The outcome of detailed studies of these species may either be a single ‘species complex’ or a group of similar species. In either case, species descriptions and distribution maps artificially simplify the reality of the complex.
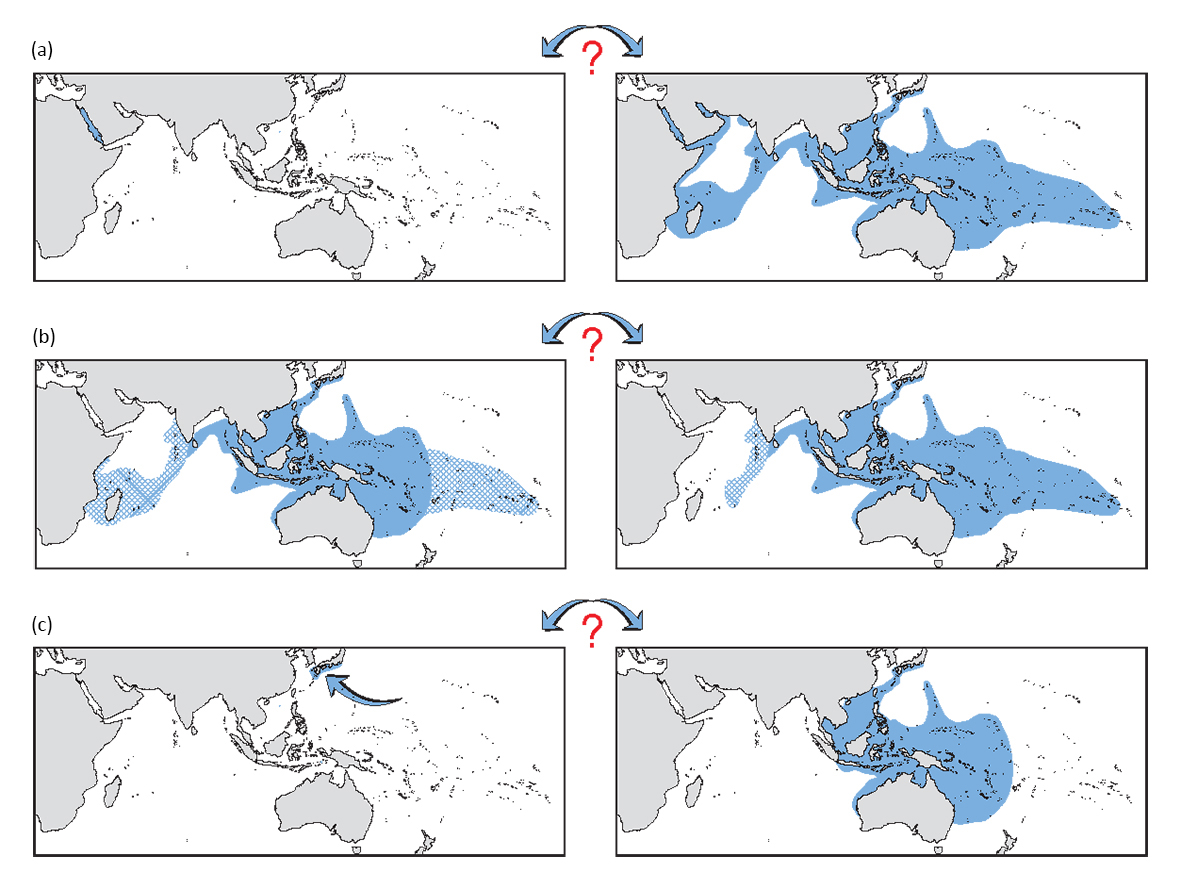
Common types of arbitrary taxonomic decisions and the geographic outcomes they represent.
- (a) Top maps. Most corals of the Red Sea are, to a variable degree, distinct from their central Indo-Pacific counterparts. In some cases the Red Sea part of this continuum is given individual species status (left map), in others (depending on level of discontinuity) the whole continuum is considered to be one species (right map).
- (b) Middle maps. Many corals of the western Indian Ocean or central Pacific are distinct from their central Indo-Pacific counterparts although there are no discrete zones of morphological or geographic separation. Both the western Indian Ocean and central Pacific parts of the continuum may be included in the species, but with doubt (left map). Alternatively, part of the range is attributed to another species (not illustrated) and the remainder may form a more cohesive unit (right map).
- (c) Bottom maps. In a relatively narrow geographic context, most corals that occur on rocky foreshores in subtropical regions are distinct from their tropical counterparts although there may be no discrete geographic boundaries to that distinction. The subtropical part of the continuum may be given individual species status (left map), or the whole continuum considered one species (right map).
Geographic patterns and synonymies
One of the principal outcomes of any taxonomic revision is the formulation of ‘synonymies’. A synonymy is usually a list of names that have been applied to a particular species together with the identity of the authors who used the names. With corals, synonymies typically include names used by different authors working in different countries at different times. They primarily focus on subspecies levels. These vary geographically just as species do, only more so. Therefore, the more comprehensive a synonymy is intended to be, the more likely it will be that it will include ‘fuzzy’ species boundaries and thus the more likely it will be that it includes arbitrary decisions.
General principles about species over large geographic ranges are:
- a) their geographic boundaries interact with other species
- b) their morphological boundaries interact with other species
- c) their synonymy interacts with other species
- d) there are no definable distinctions between species and subspecies taxonomic levels.
Further reading
For more detail on the issues summarised here, see Veron (1995a), Veron (2000a), and particularly Veron (2015) which can also be found as the Overview of coral taxonomy located under the main menu item Coral Taxonomy above.
