
Coral structure and growth
Coral structure and growth

The Scleractinia is one of approximately twenty-five orders of animals belonging to the Phylum Cnidaria (see 'Classification'). This phylum includes corals, soft corals, hydroids, jellyfish and sea anemones, all of which have the same general body plan. They are all symmetrical about a central axis (that is, they are radially symmetrical) and have a sac-like body cavity with only one opening, which serves as both mouth and anus. This opening is surrounded by tentacles which have stinging cells. The body wall, unlike that of any other group of animals except comb-jellies, consists of two cell layers, the epidermis and gastrodermis, separated by a jelly-like layer, the mesoglea. Two forms of this plan occur within the phylum, a polyp form which is usually sedentary and a medusa form which is usually free-swimming. One is the upside-down equivalent of the other.
For corals, the free-swimming stage is represented by the larval form, or 'planula'. Planulae develop either externally in the water column, following the release of eggs and sperm, often as 'bundles', by the spawning parents, or internally, being 'brooded' by the parent prior to release (see 'Reproduction'). In the latter case, the fully developed larva can settle to the reef and metamorphose almost immediately, within several hours of release. In the former case, planulae develop over several days, passing through various growth stages while being transported in surface currents before becoming competent to settle. In extreme cases, planulae can survive for hundreds of days in the plankton, and be dispersed thousands of kilometres from their natal reef.
Once the larva settles to the reef, it attaches itself and metamorphoses into a polyp, secreting the initial calcareous corallite. Hence, adult corals are basically anemone-like animals that secrete a skeleton. Some corals are solitary and look just like simple anemones when their tentacles are extended. Others, including most that are seen on coral reefs, are colonial. Although corals are primitive organisms, their skeletons, like those of many other primitive organisms, are often complex. Fortunately it is not necessary to understand much about this complexity in order to identify corals.
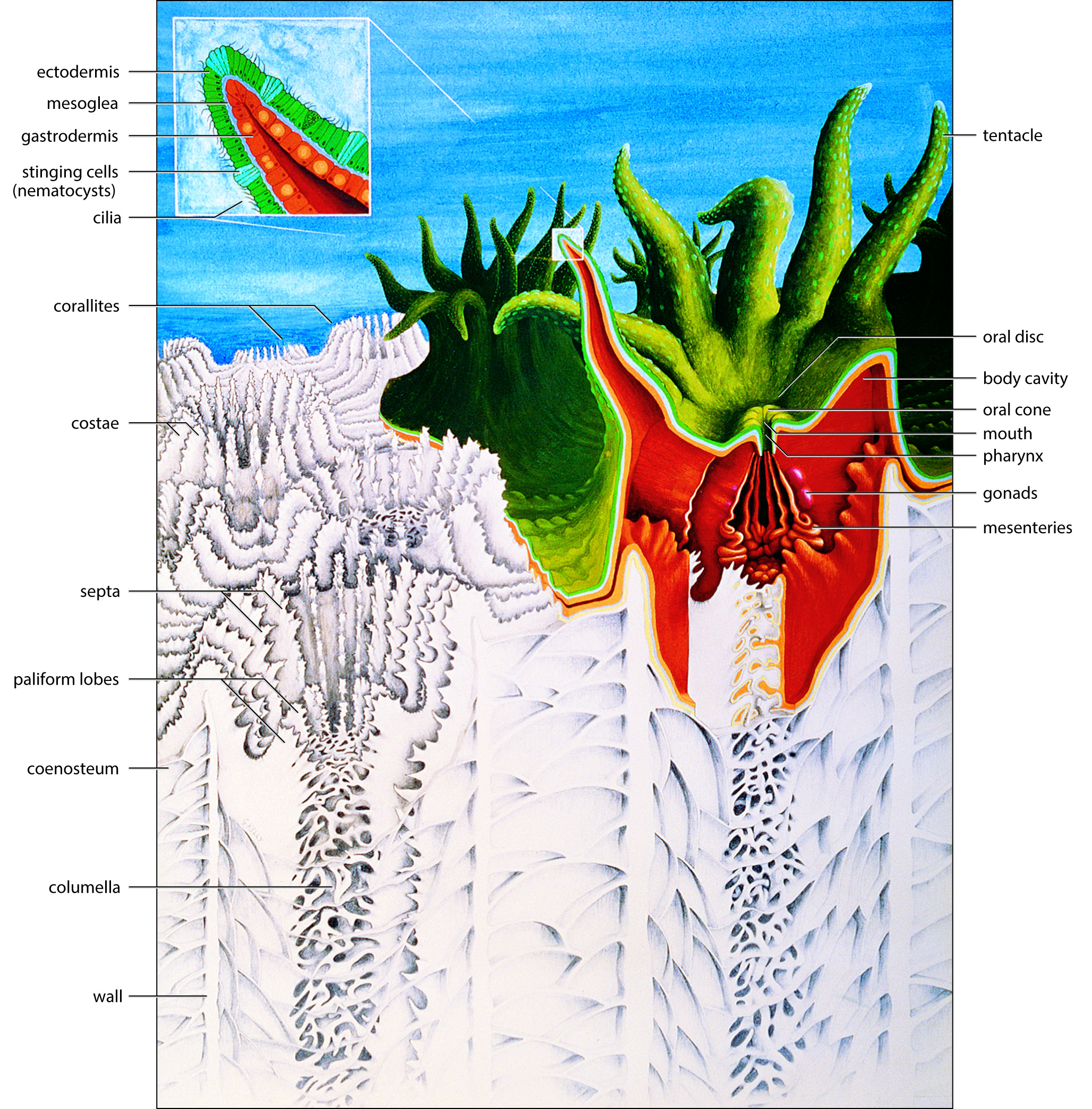
The general structure of a polyp and underlying skeleton. Painting: Geoff Kelly. Figure 1.

The polyp skeleton
The skeleton of an individual polyp, called the corallite, is a tube that contains vertical plates radiating from the centre. The tube itself is the corallite wall and the plates are the septo-costae. The tubes are joined together by horizontal plates and other structures, collectively called the coenosteum. Some polyps have an additional thin film of skeleton around the wall called the epitheca.
The wall is formed by five skeletal elements which vary in proportion in different coral families and/or genera. These elements are a) septo-costae (which become thickened within the wall), b) coenosteum (which forms a sponge-like structure), c) synapticulae (which are horizontal rods forming a lattice between the septo-costae), d) sterome (which forms a non-porous layer within the wall) and e) epitheca (which forms a thin non-porous layer on the outside of the wall). The wall is very prominent in some corals, but is inconspicuous in others where individual polyps may be indistinct.

Figure 2. The basic wall components of corals. Five examples where specific wall types are dominant. Other major families may have two equally dominant wall components: the genera Acropora, Montipora and Pocillopora have walls of mixtures of thickened septo-costae and coenosteum; the genus Heterocyathus has walls formed of mixtures of thickened septo-costae and epitheca.
a) The wall of this Acanthastrea is primarily composed of thickened septo-costae. This is also seen in some other genera, notably Favia. Photograph: Veron archives Figure 2a.

b) The wall of this Duncanopsammia is primarily composed of sponge-like coenosteum. This is also seen in the genera Turbinaria, Porites and Goniopora. Photograph: Veron archives Figure 2b.

c) The wall of this Conotrochus is partly composed of epitheca. This mostly occurs in azooxanthellate corals. Photograph: Veron archives Figure 2c.

d) The wall of this Pavona is primarily composed of horizontal rods of synapticulae. This is also seen in the genera Agaricia, Leptoseris, Psammocora and Coscinaraea, among others. Photograph: Veron archives Figure 2d.

e) The wall of this Echinophyllia is primarily composed of sterome. This is also seen in the genera Euphyllia, Galaxea, Meandrina and Pectinia among others. Photograph: Veron archives Figure 2e.
The septo-costae are the radial elements of the corallite and are divided (by the wall) into two components: the septa, which are inside the wall, and the costae, which are outside the wall. Where the wall is indistinct (as in the genera Agaricia, Pavona, Leptoseris, Coscinaraea and Psammocora) the septo-costae are single uniform elements. In solitary mushroom corals (Fungia and Cycloseris spp.) the wall is horizontally compressed, with the septa above it and the costae below it. In most corals, the septa are of different lengths and have a cyclical symmetry. They may be in cycles (usually with 6 septa in the 1st cycle, 6 in the 2nd cycle, 12 in the 3rd, 24 in the 4th and so on if present) or orders (where there is an indeterminate number of septa of each length). In practice, this cyclical arrangement is often unclear. In many corals, but especially in Turbinaria, the cyclical arrangement of septa is embellished into a pattern of fusion called Pourtàles plan, where septa of the 4th cycle curve in front of those of the 3rd cycle and fuse. This appears to be a primitive characteristic of the Scleractinia as it sporadically occurs in several families and can also be seen in the earliest fossils. The genus Porites has a unique septal plan which is used extensively in taxonomy.
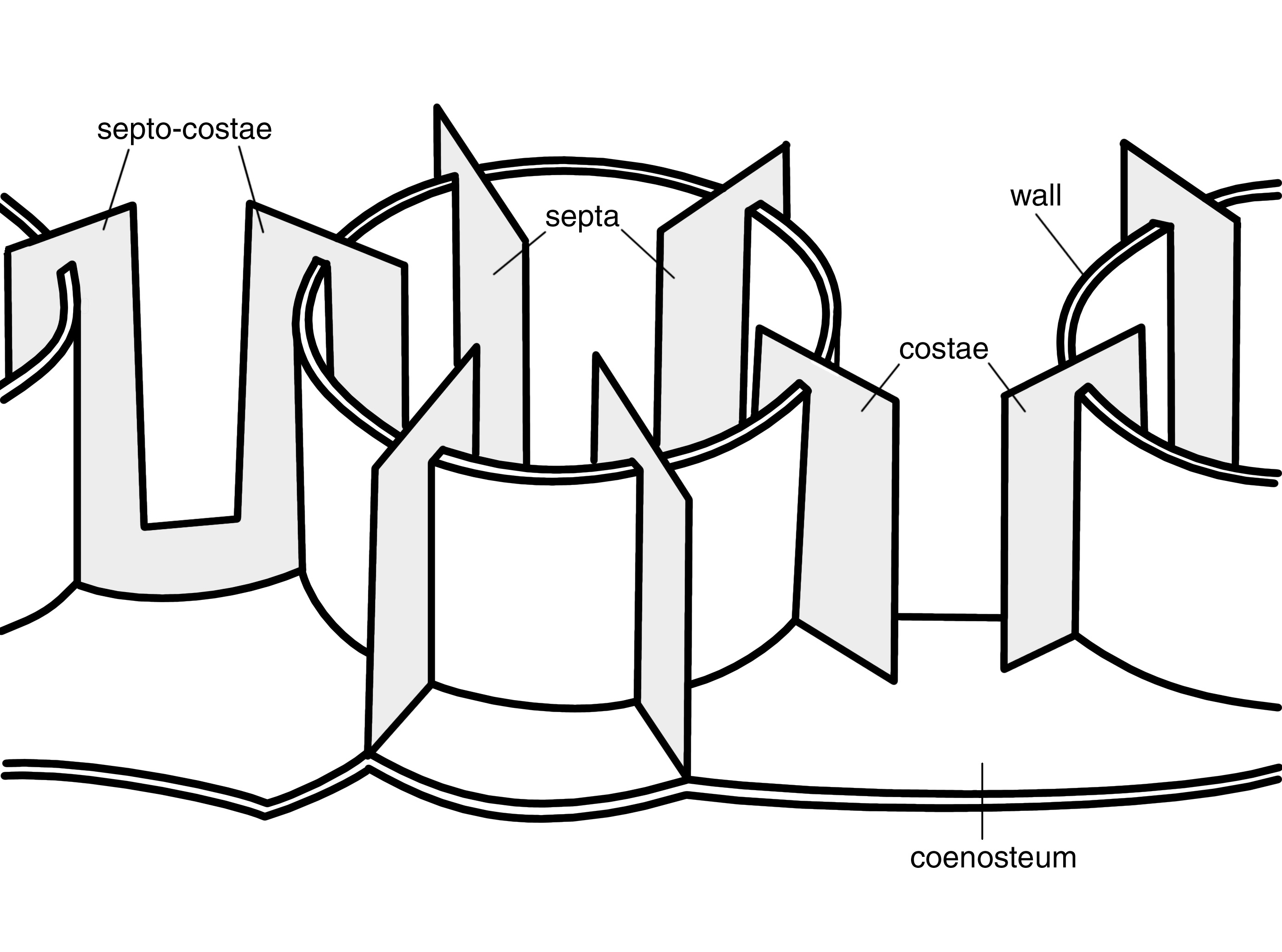
Diagrammatic representation of the basic skeletal elements of corals. Photograph: Veron archives Figure 3.
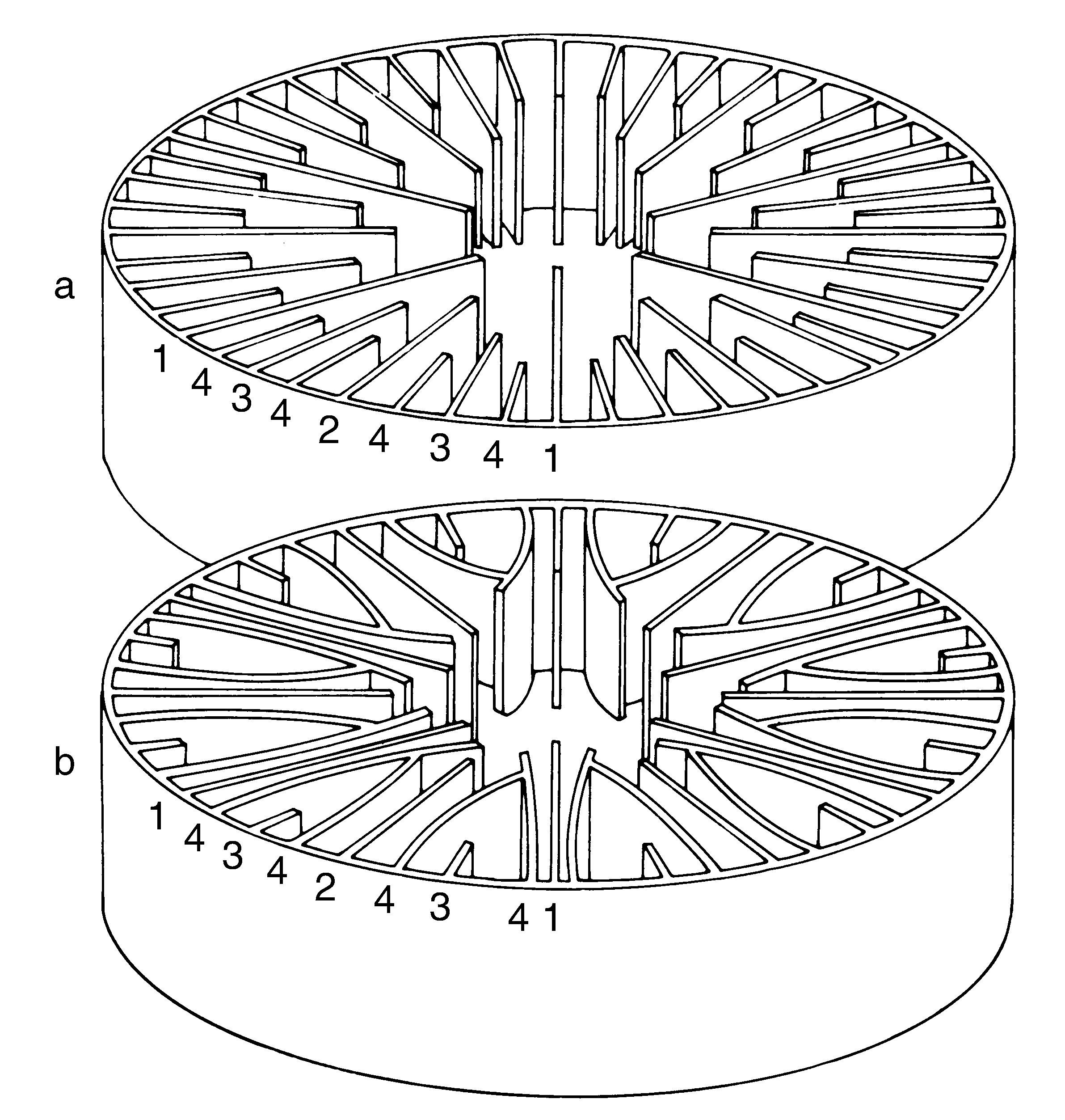
a) Normal cycles of septa. b) Pourtàles plan. Numbers indicate cycles. Photograph: Veron archives Figure 4.
Septa seldom join at the centre of the corallite (except in the Astrocoeniidae and Pocilloporidae). Instead, their inner margins usually have fine inward-projecting teeth which, in most corals, become intertwined forming a tangle called the columella. In some families, especially the Astrocoeniidae and Pocilloporidae, the columella is pillar- or dome-shaped. In others, especially the Acroporidae, it is usually absent. Many corals have pillar-like projections on the inner margin of some or all of their septa called paliform lobes and these often form a neat circle around the columella called a paliform crown. Some groups of corals have pali instead of paliform lobes. These are the result of the pourtàles plan pattern of septal fusion although the pattern may not be visible in mature corallites.

Figure 5. The appearance of the columella and paliform lobes. a) A Parascolymia showing the typical appearance of a columella composed of a tangle of spines from the inner margins of septa. Photograph: Veron archives Figure 5a.

b) A Goniastrea with paliform lobes forming a neat crown. Photograph: Veron archives Figure 5b.
There are four other parts of the skeleton which are used in general descriptions of corals (apart from being components of corallite walls as noted above): coenosteum, sterome, dissepiments and epitheca. The coenosteum is a general term for porous (not solid) skeletal material situated between the costae of corallites or between one corallite and the next. This is best seen in the genus Turbinaria where the corallite wall and the skeleton between the corallites consist of a sponge-like matrix of coenosteum. The sterome is a solid sheet which forms the inner lining of (or all of) the corallite wall. This is best seen in the genera Euphyllia, Galaxea and Meandrina, and gives the skeleton a porcelain-like finish. The dissepiments are thin, blister-like layers of skeleton which form between the corallites and are structurally similar to the sterome. The epitheca is a delicate translucent skeletal layer. It initially occurs as the basal plate deposited by the planula larva on settlement, and thereafter may continue growing to envelop individual corallites. The epitheca is always a distinct skeletal entity which is not covered by living tissue; in some genera including Astrea and Phymastrea, its growth is controlled in many colonies by tiny polychaete worms to form 'groove-and-tubercle' structures.

Figure 6. The appearance of various skeletal elements. a) Corallite walls and the skeleton between the corallites of this Turbinaria skeleton consist of a sponge-like matrix, the coenosteum. Photograph: Veron archives Figure 6a.

b) The smooth skeleton between the septa of this Catalaphyllia skeleton is the sterome. Photograph: Veron archives Figure 6b.

c) The fine blisters of skeletal material between the corallites of this Galaxea are the dissepiments. Photograph: Veron archives Figure 6c.

d) Fine skeletal structures between the corallites of this Phymastrea are called 'groove-and-tubercle' structures and are composed of epitheca. Photograph: Veron archives Figure 6d.
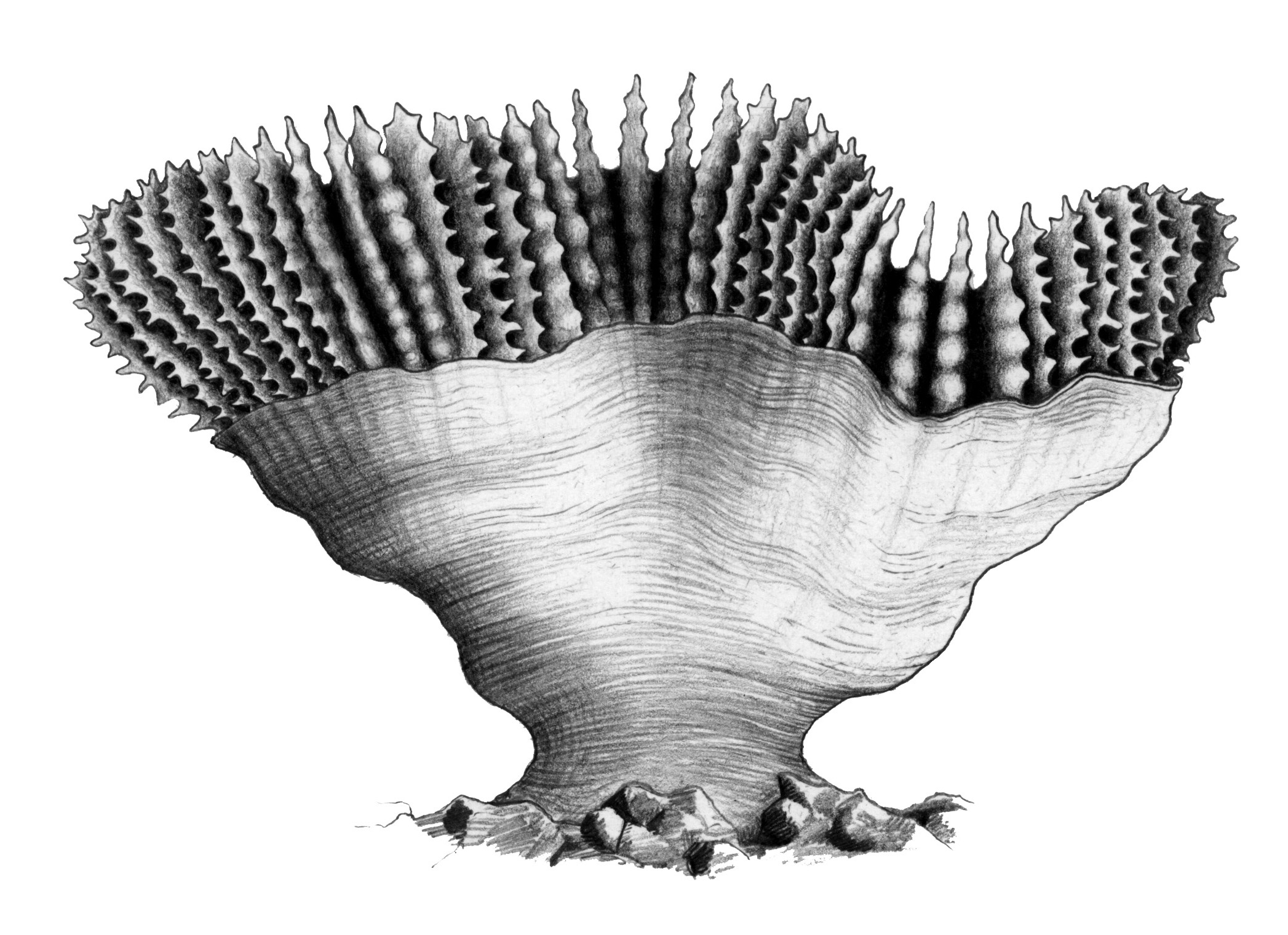
e) The skeletal layer covering the outside of this Trachyphyllia is the epitheca. Drawing: Geoff Kelly. Figure 6e.
Some skeletal structures are found only in some corals. Montipora and Porites in particular, have additional skeletal structures which are useful in identification and are discussed in sections relating to these genera. Among the other structures, monticules are primarily found in Hydnophora (where they are also known as hydnophores), but may occur in other genera. Ambulacral grooves are seen in a scattering of unrelated species.

The formation of monticules. a) Hydnophora colonies with sections of wall of variable length. Photograph: Veron archives Figure 7a.

b) Monticules where walls are as short as they are wide. The star-like structures that result are the monticules. The ribs down the sides of the monticules are costae. Photograph: Veron archives Figure 7b.

Ambulacral grooves. The grooves running along the tops of the walls are ambulacral grooves. They represent a tendency toward flabello-meandroid structure in meandroid colonies. Photograph: Veron archives Figure 8.
The polyp tissues
The coelenteron, the sac-like body cavity of the coral polyp, has a single opening to the outside. The coelenteron of one polyp is linked to those of adjacent polyps by tubes through which water circulates and nutrients are transported. The coelenteron serves many functions including digestion and the circulation of fluids for respiration and nutrition. The mouth leads to a short tube, the pharynx, which opens into the body cavity. In most corals it is short; in others (notably Goniopora and Alveopora) it is extraordinarily extendable, allowing the mouth and tentacles to protrude far beyond the skeleton to aid food capture. The coelenteron is a complex structure, made so by the skeletal structures, all of which lie outside it, but which are enveloped by it. The coelenteron is partitioned by vertical mesenteries, arranged in a radial fashion according to the position of the septa. These mesenteries give the gastrodermis a large surface area for digestion, photosynthesis and respiration, and also contain the reproductive organs. A series of coiled filaments, the mesenteric filaments, are packed along the inner margins of the mesenteries. These further extend the surface area of the mesenteries and are extruded through the mouth in response to stress.
The body wall, as with all coelenterates, is primarily composed of two cell layers, the ectodermis on the outside and the gastrodermis on the inside (see Figure 1 above). These layers are separated by the mesoglea, which is initially non-cellular but which may contain a wide range of cells after initial growth. In corals with small corallites the mesoglea is microscopically thin while in others, notably the big corallite species of Lobophyllia, it may be several millimetres thick and is of tough construction.
Extended polyps have an anemone-like appearance. The mouth is usually slit-like and may be surrounded by an oral cone. The tissue between the cone or mouth may be raised into an oral disc.

Polyps of Goniopora showing slit-like mouths and well-defined oral cones. Northern Australia. Photograph: Charlie Veron. Figure 9.
Tentacles are tubular and have the same two tissue layers as the rest of the polyp so that the cavity inside them is part of the coelenteron. Tentacles are smooth in corals that feed on detritus but most have stinging cells for defence or food capture. These cells, the nematocysts, are microscopic in size, but in most corals are grouped into wart-like nematocyst batteries, which are clearly visible underwater. Nematocysts also occur on vesicles of Physogyra and Plerogyra, which are sac-like structures composed of body wall that are inflated with water when tentacles are retracted during the day. Other cells of the ectodermis secrete slimy mucus which coats the polyp and which is moved around by microscopic cilia. The mucus is used to remove sediment from the polyp surface and is also used by detritus feeders to capture food.

Nematocyst batteries on the tentacles of a large Cynarina polyp. Great Barrier Reef, Australia. Photograph: Charlie Veron. Figure 10.
Cnidaria are the simplest organisms to have discrete nervous, muscular and reproductive systems and in corals all of these are well developed. A simple nerve net composed of both ectodermal and gastrodermal cells permeates the body wall, with connections to a variety of specialised cells responsible for sensing mechanical and chemical stimuli as well as light. A muscular system, consisting of specialised cells of both ectodermal and gastrodermal origin, allows polyps to extend and retract in response to signals from the nerve net. These signals are transmitted from polyp to polyp, as seen in the progressive retraction of polyps when part of a colony is mechanically disturbed.

Polyps of a Goniopora retracting in sequence after the left side of the colony was mechanically stimulated. Norfolk Island, western Pacific. Photograph: Charlie Veron. Figure 11.
Reproductive organs develop within the mesoglea of the mesenteries once corals reach sexual maturity, typically following several years to up to a decade of growth. This happens on an annual cycle in most species in the lead-up to reproduction, after which the organs disappear, to re-form the following year. Some corals, notably Fungia and Porites, have separate male and female sexes, being gonochoric, but most are hermaphroditic. In either case, the gonads are arranged around the base of the pharynx in radial symmetry. Some hermaphrodite corals have male and female gonads on different mesenteries, in others the testes are above the ovaries on the same mesenteries and in others the testes and ovaries grow together.
The gastrodermis has an array of specialised cells for digestion. Digestion partly occurs in the body cavity and partly inside the digestive cells themselves. Nutrients are readily moved among polyps so that neighbouring polyps have a similar rate of growth and thus do not compete for space. The gastrodermis also contains the zooxanthellae, the unicellular symbiotic algae which are essential to the growth and survival of most zooxanthellate corals.
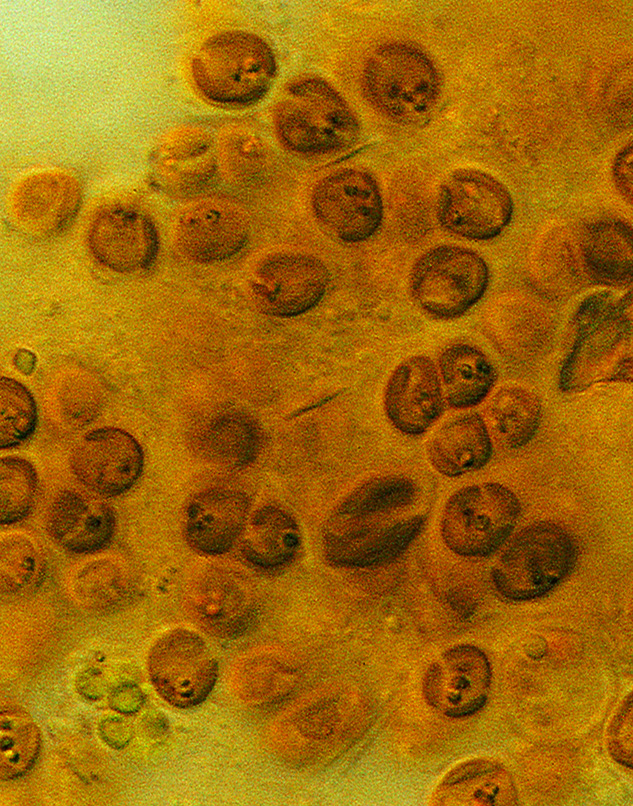
Microscopic zooxanthellae as seen when a tentacle is squashed onto a microscope slide. Photograph: Ove Hoegh-Guldberg. Figure 12.
Colony structure
Colony formation
In most corals, the overall appearance of a colony is not only determined by the way its corallites multiply. However, in some groups, the type of budding may determine the type of colony that results. In these genera, including for example Platygyra and Leptoria, the terms used to describe both budding (the formation of corallites) and growth form are usually the same. For example, the term 'meandroid' may be used to describe both the type of budding and the type of colony.
In most corals, there is a clear distinction between what is an individual and what is a colony. This is not always so, as seen in some mushroom corals, where there is a continual gradation between solitary individuals (with a single mouth) and colonies (with many mouths), as exemplified by the sequence Cycloseris › Fungia › Ctenactis › Herpolitha › Polyphyllia. In this sequence, Cycloseris and (usually) Fungia exist only as solitary individuals with a single mouth while Polyphyllia forms colonies with many mouths. A single specimen of Ctenactis or Herpolitha could be considered a solitary individual with many mouths or a colony of individuals, each with a single mouth. Likewise, in some corals there may not be a clear distinction between what is an individual and what is considered to be a row of individuals. In such cases, as exemplified by the genus Symphyllia there is a continual gradation between colonies composed of distinct polyps (corallites) to colonies where individuals are recognisable only by the existence of mouths and/or columella centres, to colonies where there is little to no sign of individuality.
All corals that form colonies do so by a process of budding, where the parent polyp divides itself into two or more daughter polyps. Intratentacular budding occurs within the tentacle ring of the parent polyp. Extratentacular budding occurs outside the tentacle ring, with daughter corallites forming on the side of the parent corallite. Some colonies, such as members of the genera Astrea and Acanthastrea, have both intra- and extratentacular buds. Another form of budding occurs in the valleys of meandroid colonies, such as some Platygyra, where individual polyps no longer have separate, complete walls.

Figure 13. Forms of budding. a) Extratentacular budding. Drawing: Geoff Kelly. Figure 13a.

b) Intratentacular budding. Drawing: Geoff Kelly. Figure 13b.
If the corallites of a colony all have their own separate walls they have exsert corallites which in some groups are known as plocoid (such as in Favia) or phaceloid (such as in Lobophyllia), depending on how elongate they are. If they share common walls they may be meandroid (such as many Platygyra) or cerioid (such as Favites), depending on whether or not they form valleys. If they are meandroid and have their own walls they are termed flabello-meandroid (such as certain Euphyllia).

Figure 14. Colony formation in corals. a) Plocoid corallites with their own separate walls. Each corallite would be associated with a single polyp in the living animal. Drawing: Geoff Kelly. Figure 14a.

b) Phaceloid colonies also have corallites with their own walls, but these are long and tubular. Drawing: Geoff Kelly. Figure 14b.

c) Corallites with common walls. Drawing: Geoff Kelly. Figure 14c.

d) Meandroid colonies have valleys holding numerous centres (each of which would be associated with a polyp in the living animal). Drawing: Geoff Kelly. Figure 14d.

e) Flabello-meandroid colonies also have valleys with numerous centres, but do not have common walls. Drawing: Geoff Kelly. Figure 14e.
Colonies which have corallites with separate walls can have both intratentacular and extratentacular budding, while those with common walls can only have intratentacular budding. Most corals have corallites or valleys that are highly interconnected with tissues linking one polyp to the next, while adjacent corallites or valleys of phaceloid and flabello-meandroid colonies may have little or no connecting tissue. The latter may compete for space and other resources, with the result that some parts of colonies overgrow other parts.
Some colonies combine two growth forms. Euphyllia and Lobophyllia colonies may be phaceloid toward the colony centre where lack of space constrains valley formation and be flabello-meandroid at the periphery where there are no such constraints. Similarly, Symphyllia colonies may have both meandroid and flabello-meandroid areas; Favia colonies may have both plocoid (separate walls) and meandroid areas; Favites and Goniastrea colonies may be both plocoid (separate walls) and cerioid (common walls). There are also many intermediate forms between plocoid and fully phaceloid and (very commonly) between cerioid and fully meandroid colonies.
A variety of other types of colony formation are found in corals, but these are uncommon.
Growth forms
The most common terms used to describe growth-form are ordinary descriptive words: encrusting (adhering as a thin veneer to the substrate); branching (forming branches); arborescent (tree-like); columnar (forming columns); laminar (plate-like - sometimes forming tiers); and so on. Sometimes, however, the terms do not have exactly the same meaning as in colloquial usage. 'Massive' does not necessarily mean large (though many 'massive' corals are large) but means solid, often hemispherical to spherical, where underlying solidity is formed by colony skeleton rather than as a veneer over the substrate. Terms such as explanate (flattened, spread out) are widely used in biology but rarely in colloquial use. Others, some of which are illustrated above such as phaceloid and flabellate, specifically deal with coral growth-forms. All terms are explained in the glossary but where their use is in Coral ID users are encouraged to give special attention to the Coral ID state definitions where the terms are defined more closely for use with this tool.
A common modification of all descriptive terms is the addition of the prefix 'sub' to the term (e.g. submassive, subcerioid, subequal), meaning ‘less than’ or ‘not quite’.

A range of typical coral growth-forms Painting: Geoff Kelly. Figure 15a.
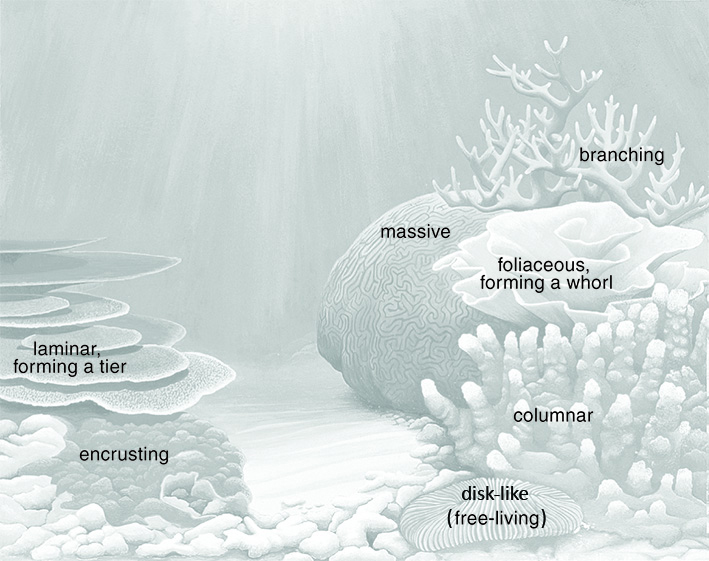
Figure 15b.
Growth rates
Growth rates, typically measured as linear extension (eg. of branches, plates or radial upwards expansion of hemispherical massive corals), range from approximately 3mm per year in the slowest growing massive corals to more than 300mm per year in the fastest branching species. Some species in the important reef-building genus Porites form large hemispherical colonies which typically grow (radially) at a rate of around 1cm per year as determined by x-rays of thin slices. Some more heavily calcified colonies of other corals, such as those of Diploastrea heliopora, grow at slower rates than this, although most are faster. Staghorn Acropora readily grows (linearly) up to about 30cm per year. Encrusting and plating corals in the genus Montipora also show rapid growth, and are among the many fast-growing species that may overtop slower-growing corals where space on the reef becomes limiting. The outcome of these inter-specific interactions, effectively slow-motion 'battles for space', between different coral species, depends on their respective arrays of offensive weaponry. Hence, growth rate, and indeed all other metabolic and reproductive processes, may be affected by disturbances that stress or injure the colony, and indeed by its location on the reef, notably in respect of illumination, water flow and turbidity (see 'Reef environments'), among other parameters. Thus, separate colonies of the same species may exhibit different rates of growth, although within the range typical of the taxon.

Growth bands in corals such as seen in this x-ray of a slice of Porites are the marine equivalent of growth rings in trees. They not only allow detailed measurements of growth rates, but can also reveal much about the environment in which the coral grew, including temperature and salinity. X-ray Photograph: Janice Lough. Figure 16.
