
Reefs
Reefs

Introduction
The term 'reef' can mean different things for different people. To seafarers reefs are solid structures that lie close enough to the sea surface to pose a risk to shipping. To most geologists and palaeontologists reefs are rock formations which have been built by organisms in the distant past but which may no longer be alive. To the biologist a reef is a veneer of living organisms forming an ecosystem which is as complex and fragile as any on Earth. These last two scientific concepts of reefs can seem as remote from each other as forests are from coal deposits, yet they share a common past. Reefs, the geological structures, are the direct products of living ecosystems and as such their formation has always been controlled by the sorts of events that control other ecosystems, both marine and terrestrial.
Even within scientific disciplines, the term 'reef' can have a wide range of meanings, a range that needs to be narrowed down to be meaningful. When considering ancient reefs, it is important to distinguish 'reefs' from the 'coral reefs' of today. Corals were not the main builders of all reefs: many ancient reefs, especially those of the Palaeozoic, were not built just by corals, but by a wide array of other organisms including algae, sponges and molluscs.
Because of the often massive accumulations of limestone, reefs provide a good fossil record from which to reconstruct their long history. Even so, the record is far from perfect, altered to greater or lesser degree by subsequent geo-tectonic processes, and by the differential fossilization of the constituent organisms.

Palaeozoic reefs
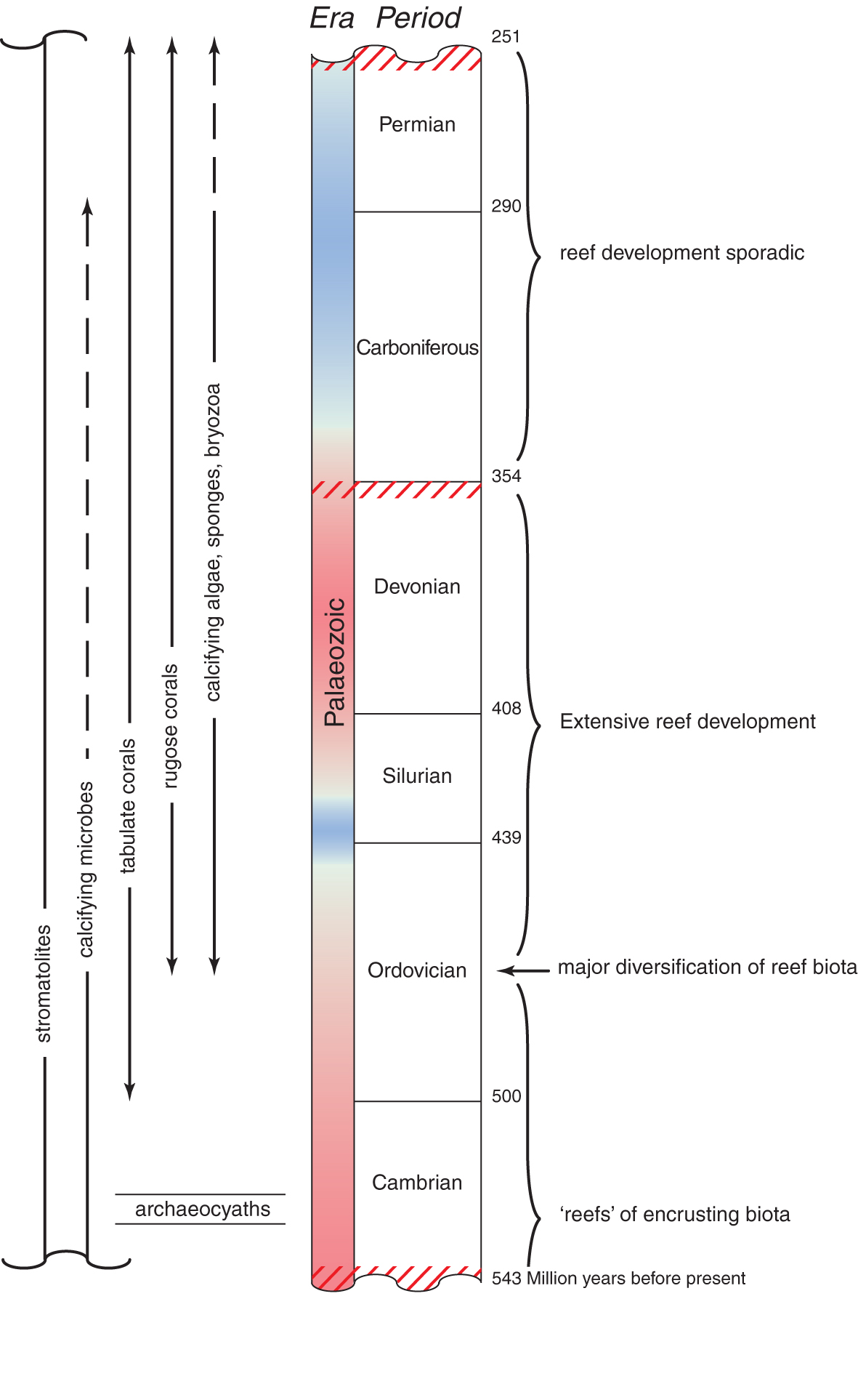
The Palaeozoic Era. Summary of reef development before the evolution of the Scleractinia. Time is in millions of years before present. Colours of the column indicate major cycles of global warming (red) and cooling (blue). Major mass extinctions are indicated by red cross-hatches. The geological longevity of principal groups of reef-building organisms are indicated on the left. Principal geological events in reef building are summarised on the right. From Veron (2000a) Corals of the World. Figure 1.
Reef-like carbonate structures of one form or another have existed on earth for at least 2.7 billion years, and hence are among the earliest evidence of life on Earth. The first reef-like limestone accumulations (excluding microbial carbonates), found in Proterozoic rocks the world over, were simple structures formed by stromatolites, hemispherical mounds of what were probably blue-green algae that entrapped fine sediment much as stromatolites do today. The first reef-like structures of animal origin were built by archaeocyath sponges of the Lower Cambrian 530-520 million years ago (mya). What reef-building there was after the extinction of the archaeocyaths was mostly due to cyanobacteria, stromatolites and some coral-like Anthozoa, all growing in shallow protected waters and containing abundant trilobites together with a wide diversity of molluscs.
Late Cambrian ecosystems persisted into the Middle Ordovician some 460 mya, when complex algae and invertebrate reef communities became widespread and reef biota greatly diversified. Stromatoporoid sponges and tabulate corals radiated at this time. Rugose corals first appeared in the Middle Ordovician and rapidly increased in number and diversity. Thus, algal communities were largely replaced by communities of skeletonised metazoans. By Late Ordovician some 450 mya, colonial rugose and tabulate corals had greatly diversified in shallow water and formed coral patch reefs, along with stromatoporoids, other sponges and calcareous red algae. These reefs had little wave resistance and did not form solid platforms although stromatoporoids and tabulate corals formed massive colonies several metres in diameter. These are the oldest known reef coral communities and were possibly the outcome of symbiotic animal/algal associations. For at least 150 million years different combinations of these algae, sponges and corals built reefs around the tropical world. Following the Ordovician-Silurian mass extinctions, Silurian reefs became abundant and diverse from ca 435 mya, and some reached massive proportions, the first truly wave-resistant carbonate platforms.

Living stromatolites in shallow water. Shark Bay, Western Australia. Photograph: Paul Copper. Figure 2.
In the Middle to Late Devonian some 370 mya, reefs reached maximum development for the Palaeozoic. What remain today of these reefs are sometimes of enormous size and have the full range of structural characteristics of modern reefs. They had a high diversity of frame builders including microbialites, calcifying cyanobacteria, stomatoporoid and other sponges as well as tabulate and rugose corals. They also contained a diversity of other fauna and flora, perhaps comparable in diversity to modern reefs.
Latest Devonian reefs are relatively rare and throughout the entire Carboniferous most reef-like constructions were relegated to relatively deep water. This was an interval of great climatic upheavals, sea-level fluctuation, and low ocean temperatures, all probably contributing to environments hostile to reef development. During the Late Carboniferous to Early Permian, phylloid (leaf-like) algae so completely dominated would-be reef substrates that most reef biota may have been displaced. Late Permian reef-like structures were characterized by frondose bryozoans, a wide range of sponges, algae, and encrusting tube-like Tubiphytes (of unknown taxonomic affinity), all with little cementation. Corals were uncommon everywhere.

Silurian Reef. Greenland. Photograph: Paul Copper. Figure 3.
The causes of the end-Permian extinctions some 252 mya are elusive, but as far as reef biota are concerned they were probably the result of ocean acidification and anoxia due to extensive volcanism and the massive release of methane clathrates off continental shelves. It has been estimated that 80-95% of all species became extinct. Among these were all corals.

Devonian Reef. North-western Australia. Photograph: Charlie Veron. Figure 4.
Mesozoic and Coenozoic fossil reefs
No reef building occurred for 12-14 million years after the end-Permian mass extinctions. Only stromatolites remained common and it was not until the Middle Triassic that reef-like structures first re-appeared, these having a fauna dominated by sponges, algae, bryozoa and Tubiphytes, much like those of the Late Permian. However, by Late Triassic massive scleractinian reefs were abundant, especially in the Tethys Sea, which remained a centre of reef development throughout the Mesozoic.
The end of the Triassic some 201 mya was marked with a mass extinction that was not the equal of the extinctions that marked the end of the Palaeozoic 45-50 million years earlier, but it may have rivalled the extinctions at the end of the Mesozoic. The main impact on reefs was the collapse of the reefs of the Tethys, perhaps because of anoxic conditions correlated with sea-level and climatic changes. Once again massive volcanism, from rifting of Pangea, with resulting greenhouse gas emissions, appears to be implicated, yet whatever the cause, reef building did not recover for 6-8 million years. The inheritance of the Jurassic was a remnant of these extinctions, a depauperate although diverse suite of scleractinian genera. Early Jurassic reefs are rare everywhere in the world and all Triassic genera had become extinct by then.
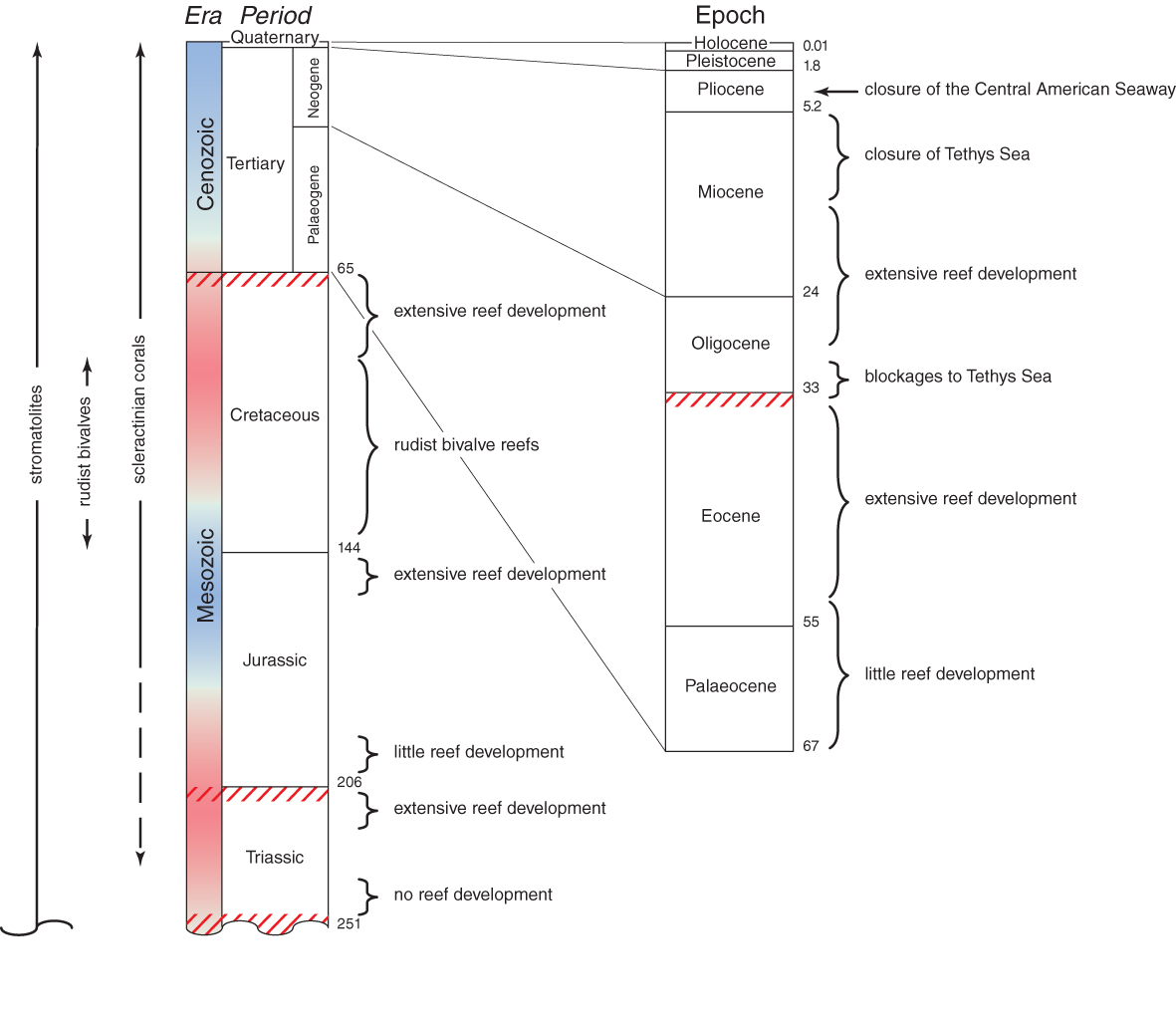
Mesozoic and Coenozoic Eras. Summary of reef development during the evolution of the Scleractinia. Colours of the column indicate major cycles of global warming (red) and cooling (blue). Major mass extinctions are indicated by red cross-hatches. The geological longevity of principal groups of reef-building organisms are indicated on the left. Principal geological events in reef building are summarised on the right. From Veron (2000a) Corals of the World. Figure 5.
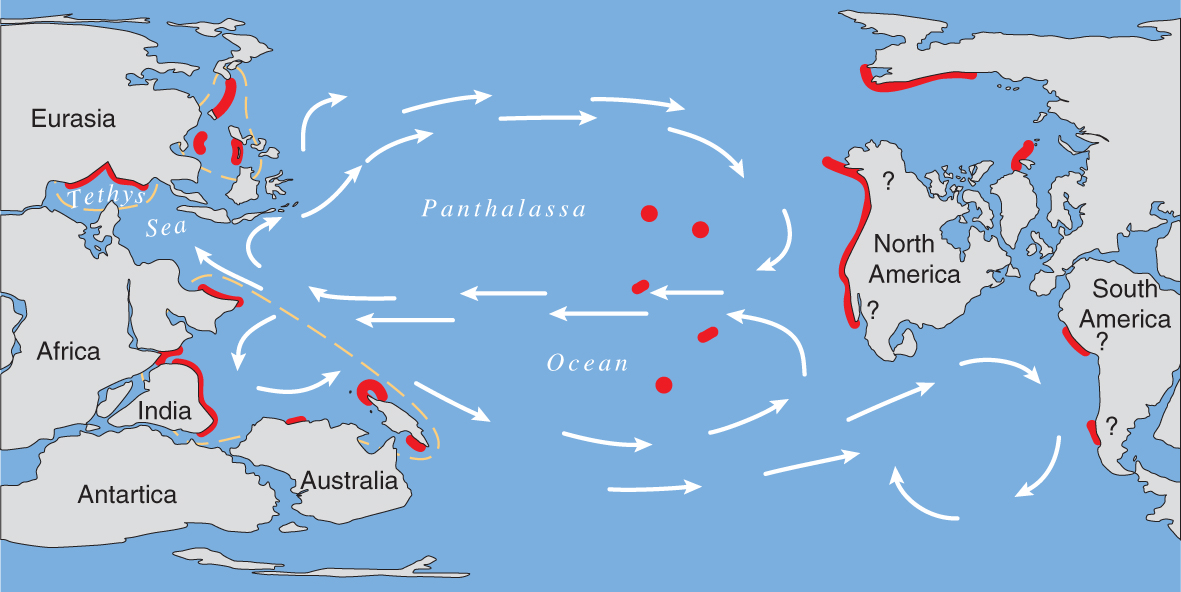
The Late Triassic showing the distribution maximum of coral reefs. From Veron (2000a) Corals of the World. Figure 6.

A Late Triassic reef.. Dolomite Ranges, Italy. Photograph: Michael Hautmann. Figure 7.
The Jurassic recovery was slow but by Middle Jurassic reef development had again proliferated in the Tethys of Europe and in the Mediterranean. Complex ecosystems underwent a major radiation, enhanced by a warming climate and extensive flooding of shallow shelf areas. Elsewhere reefs remained poorly developed, especially in the Panthalassa and thus it may have remained throughout most of the Jurassic.
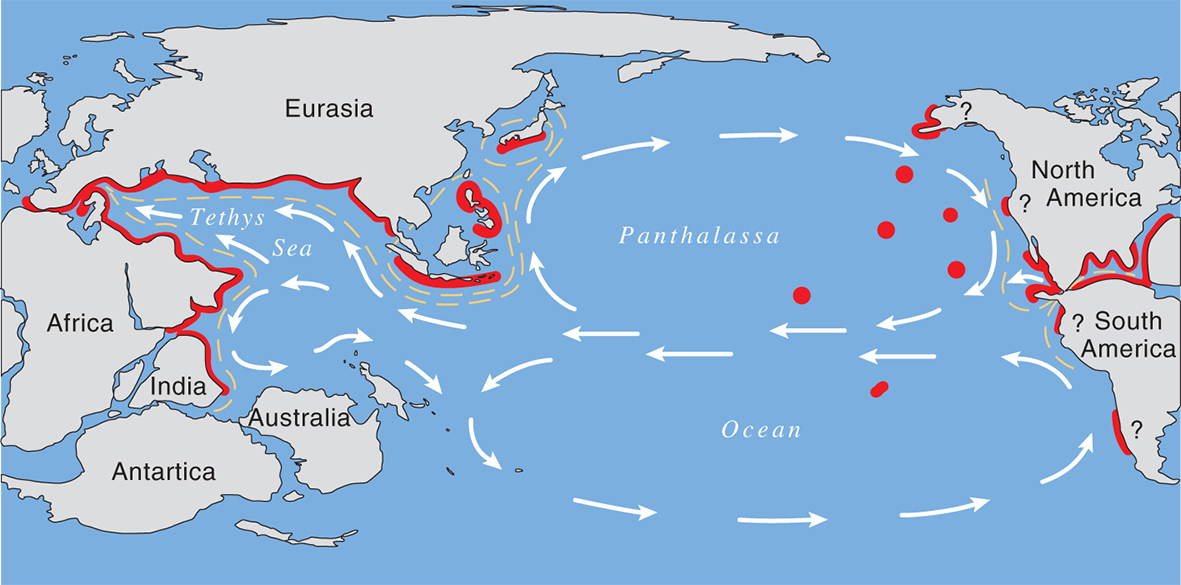
The Late Jurassic showing the distribution maximum of coral reefs. From Veron (2000a) Corals of the World. Figure 8.

Jurassic calcareous ranges. Austria. Photograph: Harald Krenn. Figure 9.
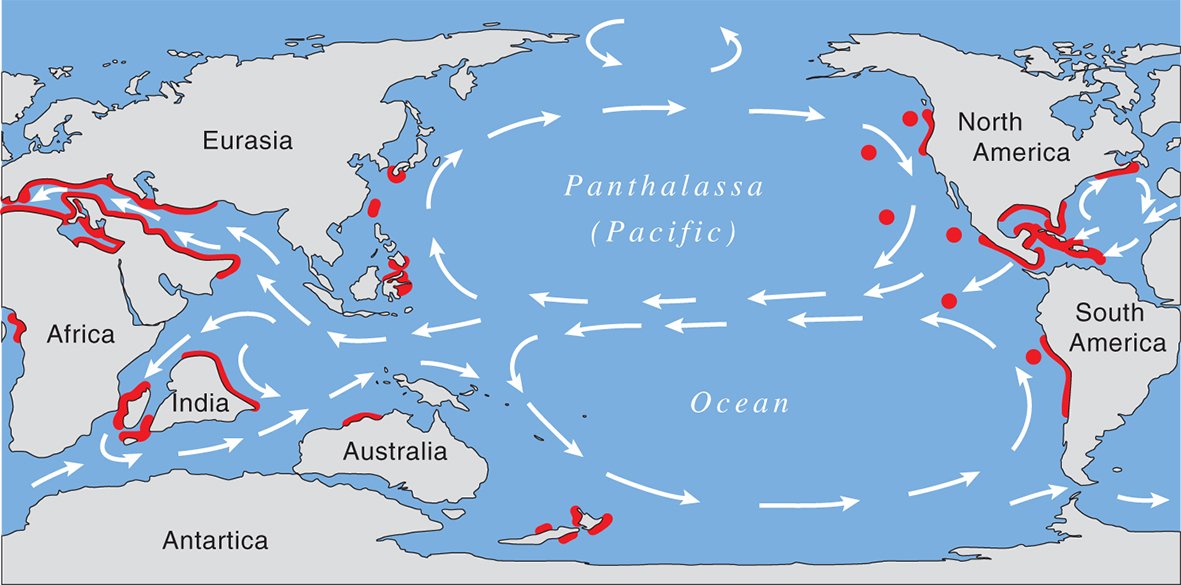
The Late Cretaceous showing the distribution maximum of coral reefs. From Veron (2000a) Corals of the World. Figure 10.
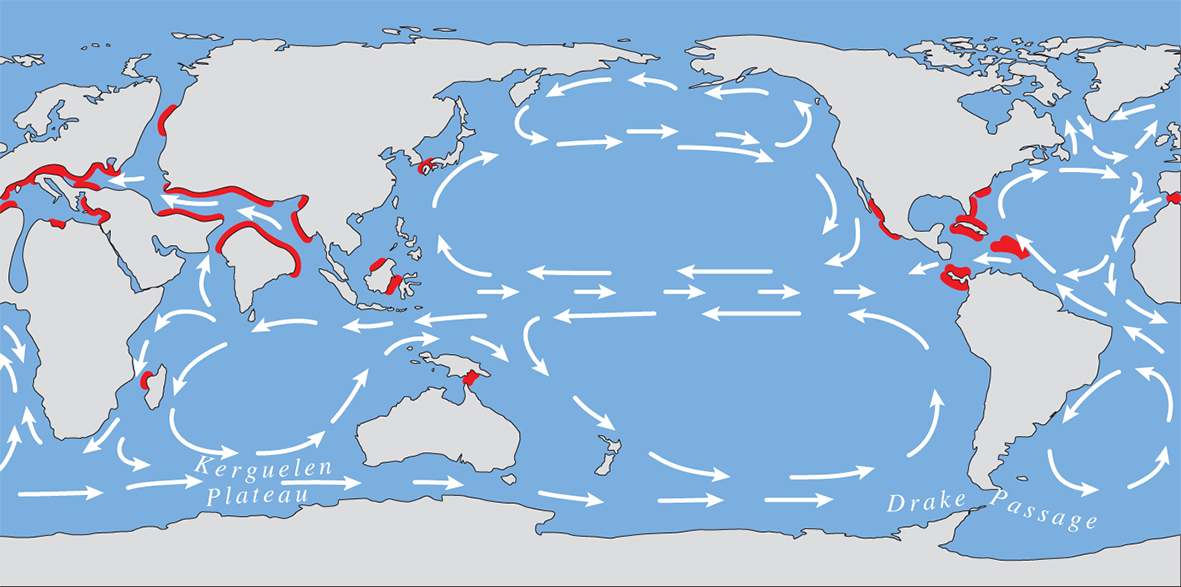
The Late Eocene showing the distribution maximum of coral reefs. From Veron (2000a) Corals of the World. Figure 11.
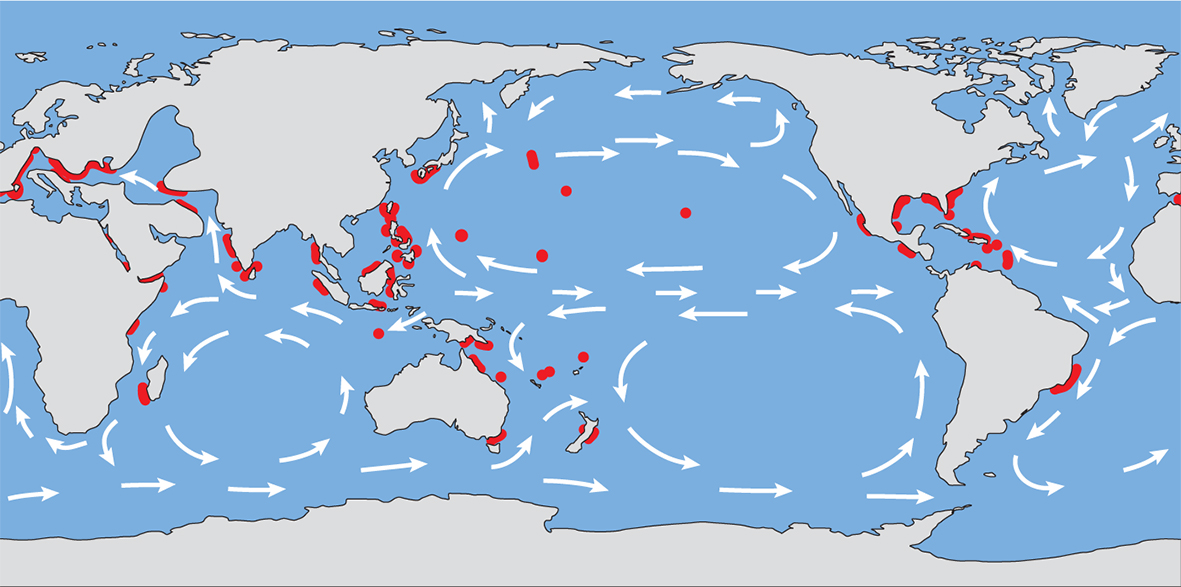
The Late Miocene showing the distribution maximum of coral reefs. From Veron (2000a) Corals of the World. Figure 12.
The beginning of the Cretaceous some 145 mya was not marked by any extinction event that had a major impact on reefs which continued to thrive throughout the Early Cretaceous. There was, however, an increasingly drastic change in coral communities over this time. Rudist bivalves, a previously obscure group of molluscs, progressively displaced corals as the dominant reef biota, and thus it remained for 30 million years. During this interval, zooxanthellate corals coexisted with rudists, but largely in separate, slightly deeper habitats. The reefs of that time probably resembled inshore fringing reefs of today, repeatedly destroyed by changing sea levels and consisting mostly of banks of entrapped sediment with no algal cementation. Rudists were probably zooxanthellate, or hosted other similarly-functioning endo-symbionts and, as they had a lesser amount of aragonite in their shells than corals, probably survived acidic conditions better than the corals.
Repeated environmental upheavals probably had a greater influence on evolutionary change in the Cretaceous than did continental movements. The Late Cretaceous was a time of extreme sea level change, periodically flooding up to 40% of the continents and creating a Super-Tethys Ocean over much of present-day Europe. Consequences for reefs are unknown, but the continually changing sea levels of the Late Cretaceous would have had a great impact on coral communities.
Much of the Middle Cretaceous was characterised by release of carbon dioxide due to extensive volcanism around the continental plate margins and this, together with the accumulation of organic matter associated with sea level changes, may have increased the acidity of much of the ocean surface. Ocean and atmospheric temperatures over a range of latitudes from the equator to the poles were much higher than they are now. This would have varied greatly over time, but subtropical conditions may have periodically extended to 45°N and possibly 70°S, and there were no polar ice caps. These conditions would have resulted in weaker ocean currents than we have today. Corals would have been far more widely dispersed and there would have been a much greater development of distinctive regional provinces.
By the close of the Mesozoic, the flooding of the continents had ceased and the warm climates that had dominated the Cretaceous had begun a long and irregular decline towards a glacial mode. Reefs did not proliferate anywhere for long after the end-Cretaceous extinctions 65 mya. Eocene reefs are mostly poorly preserved, but were extensive in area. An end-Eocene extinction event some 34 mya, combined with progressive blockages of the Tethys Sea in the Early Oligocene created a hiatus in reef development, but did not greatly diminish coral diversity.
There were two post-Miocene events which greatly affected reef development. The first was the closure of the Central American Seaway, finally separating the faunas of the Indo-Pacific and Caribbean some 3 mya. The second was the Pleistocene glaciations which, because of low sea temperatures and episodes of freshwater inundation stopped reef development in the far eastern Pacific, much of the Caribbean as well as high latitude locations of the western Pacific. More importantly, low sea levels aerially exposed all coral reefs repeatedly.
Modern coral reefs
Modern reefs are not necessarily coral-dominated. Some, forming intertidal structures in the Caribbean and Bermuda, are made by vermetid 'worms' (molluscs), and others in the Red Sea and elsewhere are built predominantly by coralline algae. There are also widespread subtidal structures dominated by serpulid worms (annelids) and odd reefs everywhere owe their existence to other organisms, especially oysters. However, in terms of quantity, reefs dominated by organisms other than scleractinian corals and their allies (including a few small taxa such as the blue coral Heliopora and the fire coral Millepora) are insignificant and have little in common with coral reefs other than in the material of which they are made. Such structures excluded, it is still necessary to narrow the term 'coral reef', for some deep ocean corals form extensive structures, built by one type of coral (Desmophyllum pertusum), which are commonly called reefs. These have none of the characteristics of coral reefs as geological structures (they do not form solid limestone) or as biological structures (they do not depend on photosynthesis nor are they biologically diverse); they warrant a separate name, 'bioherms'.

Desmophyllum pertusum, an azooxanthellate coral that forms bioherms up to 30m high at depths of 200-1000m. Norway. Photograph: Erling Svensen. Figure 13.
Although coral reefs are principally made of calcium carbonate derived from coral, corals are incapable of building reefs on their own. Coral skeletons need to be cemented into solid rock by coralline algae. Coralline algae generally have a wider distribution range than corals, but those that cement coral debris into reefs flourish only in shallow, turbulent, well-lit environments and thus it is they as much as corals that determine where highly consolidated reefs grow. They may also have a dominant influence on how fast they grow, or if they grow at all in deeper water.
Reef and other biogenic carbonates
Although most (90-95%) of terrestrial limestone is derived from reefs built by a variety of animals in shallow marine environments, this by no means reflects the principal source of carbonates on Earth. Around 90% of all today's marine carbonates are deep-sea sediments derived primarily from plankton, mostly foraminifera and coccolithophores. Due to their deep water location these are rarely uplifted to form geological rock formations on land, rather they are either dissolved in the oceans or are subducted into the Earth's mantle. Another 5% of carbonates of mixed composition are found on continental slopes. Some of this has, over geological time, been transformed into petroleum – oil, gas and condensates. Elsewhere in a few regions, these carbonates have been consolidated and uplifted or otherwise exposed on land.
Only about 5% of all carbonates today are of coral reef origin, although this small proportion includes vast tracts of mountain slopes like the calcareous reefs of Austria, formed from millions of years of accretion. The proportion of total carbonates which are represented by living reefs is probably less than 1%.
Types of reefs
There have been many attempts to classify different types of reefs. All lack general agreement because there is continuous variation from one reef type to another, also because they can be classified according to their geological history, their shape, their position relative to land masses and by their carbonate composition. In principle, these types of classification can be merged into three broad categories: 'fringing reefs', 'barrier reefs' and 'atolls' (Darwin's original groups).

Fringing reef of a continental island. Lizard Island, Great Barrier Reef. Photograph: Charlie Veron. Figure 14.

An outer (ribbon) reef of the Great Barrier Reef. Great Barrier Reef, Australia. Photograph: Charlie Veron. Figure 15.
Fringing reefs are mostly close to coastlines, are usually unconsolidated wherever protected from wave action, and usually have a high component of non-carbonate sediment. Barrier reefs are offshore and are composed of wave-resistant consolidated limestone, usually laid down on top of bedrock of terrigenous origin. Atolls are usually (but not necessarily) a wall of reefs enclosing a central lagoon, formed on top of old, subsided islands, typically of volcanic origin. Charles Darwin was the first to comprehend how atolls formed, documenting and illustrating the process in his 1842 book (Darwin, 1842). Atolls are initiated as a series of fringing reefs surrounding an island. These gradually becoming a barrier reef enclosing a shallow lagoon as the island erodes and subsides. An atoll subsequently forms when the island vanishes beneath the ocean surface. The Tahitian archipelago today provides examples of all states in this process. As the shape of both barrier reefs and atolls is largely determined by the bathymetry of the substratum, there are all manner of intergrades between them. Likewise, 'fringing reefs' gradually become 'barrier reefs' with increasing distance from the shore. Reefs which do not conform to any of these descriptions are commonly called 'platform' reefs. These typically occur in relatively shallow waters (< 100m depth) on continental shelves, and are a major feature of the mid-shelf region of Australia’s Great Barrier Reef (GBR). Platform reefs of the GBR are typically distributed between the nearshore reefs that fringe continental islands and, rarely, the mainland shore, and the offshore ‘outer barrier reefs’ located near the edge of the continental slope, of which the ‘Ribbon Reefs’ are the most well-known.

The edge of a platform reef. Great Barrier Reef, Australia. Photograph: Charlie Veron. Figure 16.
Distribution of reefs
The reefs and coral faunas of geological intervals are a combination of inheritance from the past and the outcome of more contemporary events. The distribution of modern reefs are an inheritance of the Tethys-dominated world of the Eocene, subsequently modified by the obliteration of both the Tethys Seaway (Miocene) and the Central American Seaway (Pliocene), see above. Today, the global area of coral reefs is estimated at some 255,000 square km, accounting for less than 1 % of the ocean surface. The present estimate is a significant reduction from earlier estimates of contemporary reef area of ca 600,000 sq. km. Although presently unquantified, there is also a large global ‘inter-reefal’ area, ranging between 30m and 150m depth — the ‘mesophotic zone’ (see Mesophotic environments) — which also supports reef-building corals.
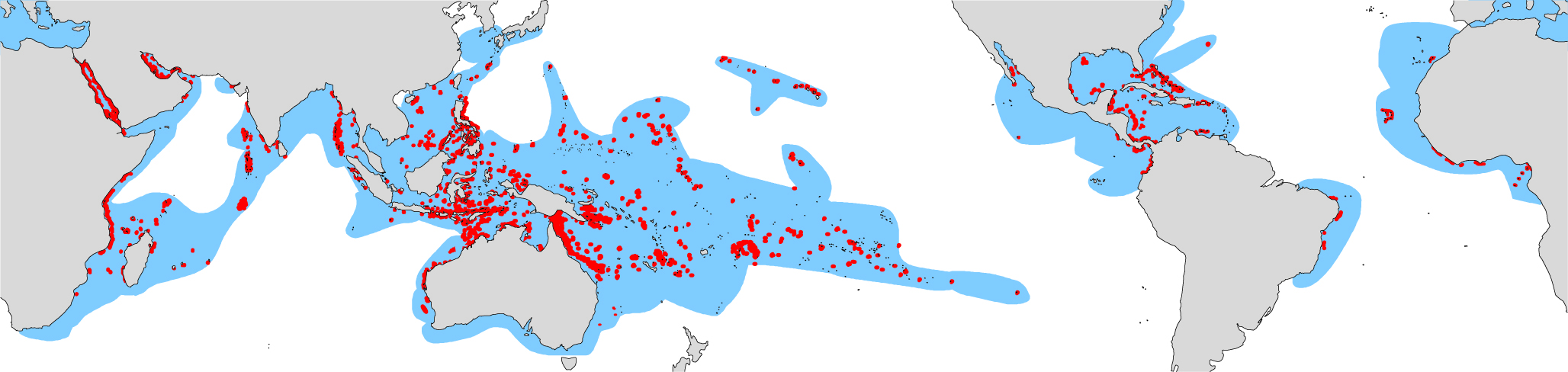
Distribution of the world’s coral reefs (red). Blue areas indicate areas in which zooxanthellate corals occur. From Veron (2000a) Corals of the World. Figure 17.
The Indonesian/Philippines archipelago has the world’s greatest concentration of reefs and the greatest coral diversity. Other areas of reef concentration are the Great Barrier Reef of Australia, the Red Sea and the Caribbean, the latter having a much lower diversity than all major Indo-Pacific regions. Large oceanic reef systems occur across the western – central Pacific, including Micronesia, Melanesia and Polynesia, and also in the Indian Ocean, notably the Maldives, Lakshadweep, Chagos and Mascarene Archipelagos. Other than the reef systems of the Caribbean Sea, the Atlantic has far fewer oceanic reefs than the other two tropical oceans.
Reefs and coral distributions
It is true, with rare exceptions that without corals there can be no coral reefs, however it is not true that without reefs there are no corals. Rather the opposite: there are at least as many corals (excluding azooxanthellate species) growing in environments that will not support reef development as there are in environments that will. This is because the constraints of temperature, light availability, aragonite saturation state, sediment load, depth and (lack of) turbulence may act independently or in various combinations to prevent reef growth (see Kleypas, McManus and Menez, 1999). These environmental constraints clearly interlink to produce habitats in which corals might flourish yet do not produce skeletons quickly enough to overcome erosion, or where encrusting coralline algae do not grow. Without the cementing action of algae, coral debris remains unconsolidated and can be swept away by storm waves. Unconsolidated rubble accumulations occur primarily where conditions for coralline algae growth are poor – where wave action is weak, the water turbid, or the temperature frequently falls below 18°C. Conditions for coral growth, however, may be good in such places. Many places where corals are abundant illustrate this; for example, the sea off the northern coast of central Australia is shallow and moderately turbid. There are no reefs, yet 60% of all Great Barrier Reef species grow there, some forming colonies as big and healthy as those on any reef. Long-term ecological issues aside, almost all species of Indo-Pacific corals grow as well in non-reef habitats as they do on reefs. For a few species, larger populations occur in inter-reefal habitats than on the reefs themselves.
Reef growth and coral diversity

Rich reef with low diversity. Reefs can be well developed in low diversity areas. In this case the entire reef is composed of fewer than 10 species. Clipperton Atoll, eastern central Pacific. Photograph: Charlie Veron. Figure 18.
Globally, coral diversity is highest in the Coral Triangle (see 'The Global Pattern' diversity map in the Overview of Coral Distributions), and within this region, it peaks in areas of locally-high habitat diversity, where various reef types and a broad range of environmental conditions coincide to provide a wide array of different niches for coral growth. Each coral species has a particular set of life history traits that determine specific habitat preferences, and hence the larger the local variety of niches, the higher the coral diversity. Outside the CT, this general rule still applies, although overall diversity declines both latitudinally and longitudinally, particularly in the subtropics and eastwards across the central and eastern Pacific.
There is, however, a widely held misconception that reef growth is somehow dependent on coral diversity, probably stemming from the observation that most reefs occur where coral diversity is at least moderately high. However, reefs of the Caribbean have a lower diversity than most Indo-Pacific reefs, and well-formed reefs of the Abrolhos Islands of Brazil have less than 20 species. Clipperton Atoll in the far eastern Pacific has actively growing reefs which provide significant habitat diversity yet only seven species of coral occur there, just three of which contribute to reef-building.
Controls on reef development
Reef-building corals and the reefs that they build have similar, tropical, distributions. This creates an interesting dilemma: which comes first, the coral or the reef, or at least the antecedent topography on which corals initially settle and grow? Corals require hard substrate on which the planula larvae settle from the plankton, metamorphose and attach, prior to colony formation. This antecedent topography typically includes the submerged sides of continental and oceanic islands, shallow submerged banks or shoals of terrestrial or biogenic origin, the latter including fossil reefs created during previous periods of reef-building. Reefs on continental mainland shores are best developed away from the influx of freshwaters in coastal runoff. Examples include the extensive fringing reef systems of the northern and central Red Sea and the Ningaloo Reef tract of Western Australia.
Substrate availability, sedimentary regimes, bathymetry, tidal regimes, turbulence, water quality, nutrients and biotic factors all affect reef development by controlling coral growth. These operate on local scales in shallow tropical seas primarily by affecting substrate conditions and light availability. On larger scales of space and time, sea levels and temperature, light and aragonite saturation, are the most important limiting environmental parameters.
Sea Level
Because coral reefs built by zooxanthellate corals can exist only in very shallow water, they are continually affected by sea-level changes due to the land or sea rising or falling. Darwin recognised this long before the phenomenon of changing sea levels per se was actually discovered, yet even now it is not generally appreciated that coral reefs have not always looked like they do today. In times past, sea-level changes have been so rapid and so frequent that what are reefs today would have alternated between being exposed limestone islands and submerged limestone pinnacles. This transition occurred at least eight times during the Pleistocene, in response to repeated glacial and inter-glacial periods. The strongly zoned reefs of today (see later) are artifacts of an unusually long period of relatively stable sea level, the Holocene high-stand.
Temperature
There is no correlation between coral diversity and reef formation (see above), but there is a clear correlation between ocean temperature and zooxanthellate coral distribution and also between temperature and formation of highly consolidated reefs. Reefs only occur where sea temperature does not fall below 18°C for extended periods of time, however approximately half of all coral species occur where temperatures regularly fall to 14°C. This appears to be a contradiction, but the likely solution is that temperature controls the formation of reefs through ecological processes. Reef building requires carbonate production and that requires coral-dominated ecosystems, along with the ‘cementing’ coralline algae. In the tropics, coral ecosystems exist because they are able to out-compete algae-dominated ecosystems, but this is not the case in higher latitudes. Thus, the highest latitude coral reefs in the world all have mixtures of coral and algal communities which compete, and the corals struggle to get enough light and space to survive.
Ecology and biology of reef building
If corals grow in sufficient quantity and the rate of both coral calcification and algal cementation exceeds that of erosion, the resulting accumulation of calcium carbonate can form limestone reefs.
The success of reef-building depends on speed, which is aided for reef-building corals by the symbiotic zooxanthellae which harness the energy of the sun to help power the process. Why do these organisms invest so much energy into building something that is dead? One aspect is that skeletons are needed to form colonies and colonies are needed to build large wave-resistant reefs.
The two evolutionary innovations of colony formation and algal symbiosis clearly go together and have very likely evolved together. The importance of this is demonstrated by the fact that most living Cnidaria that reap the advantages of reef-building are both colonial and symbiotic. The two major groups of extinct reef-building corals (rugose and tabulate corals) were also colonial, however it is not known if they were also symbiotic.
Corals are the most prominent reef-builders today because, due to symbiotic zooxanthellae, they can harness the energy of the sun to make building material sufficiently quickly that the rate of reef accretion outstrips that of erosion. However, not all zooxanthellate corals contribute to reef-building; perhaps half of all species do not grow in environments suitable for reef growth, especially where the water is too turbid or too cold or where there is limited hard substrate, and others have skeletons which are fragile and easily removed by wave action.
Symbiosis Reefs of today are an ancient phenomenon without parallel elsewhere in Nature – a specific result of a symbiotic relationship between plants and animals. Just what sort of plants and animals is not critical, but the relationship between them is. Symbiosis allows the effectively limitless resources of sunlight and seawater carbonates to be harnessed and used to construct an ecosystem. In effect, symbiosis allows reefs to be built by animals because it gives them the energy-generating capacity of plants. What the animal gives the plant is a medium in which to live that is stable yet exposed to sunlight. What the plant gives the animal is an enhanced capacity to remove metabolic waste, an enhanced capacity to calcify, a direct nutrient source and a capacity to concentrate and re-cycle limited nutrients including nitrates and phosphates. Symbiosis thus allows corals to exist in an almost nutrient-free environment, including the nutrient deserts of the open oceans.
There are few non-symbiotic scleractinian corals that live exclusively or preferentially on reefs, the genus Tubastrea being perhaps the best known exception, with the vast majority of these ‘ahermatypes’ occurring in deep water habitats. It is an interesting fact that despite not hosting zooxanthellae, at least one species of Tubastrea calcifies at a similar rate to a zooxanthellate species (Galaxea fascicularis) (Marshall (1996)). However, there are so few azooxanthellate coral species on reefs that these make an insignificant contribution to reef building.
Clearly, the evolutionary advantage of symbiosis is very great, but the evolutionary cost is also great. Zooxanthellate corals are constrained to live near the ocean surface, the most hostile of marine environments both physically and biologically. Most importantly, symbiosis constrains corals to live in places where they must compete with macroalgae.
Reef growth rates
Speed of coral growth is critical to both coral survival and reef-building once basic environmental needs are satisfied. This is because corals and coralline algae must build reefs fast enough to outstrip the rate of erosion.
Rates of reef growth (accretion) can be measured directly from cores taken from reefs or by a wide range of indirect measures of carbonate production. Normally, the maximum rate of reef growth is about 0.6 metres per century although rates in optimal conditions may reach three times this. These optimal conditions only occur where the water is shallow and clear and currents are strong, the probable reason why continuous areas of reefs usually occur where the tidal range is great and the ocean floor is shallow. These environments provide high light levels combined with continuous flushing and nutrient transport.
Rates of reef erosion determine the appearance and existence of reefs every bit as much as rates of growth because accretion must out-strip erosion for any reef to form and survive. There are few reliable estimates of rates of reef erosion because they are too slow to be directly measurable, however best estimates suggest that around 90% of all calcium carbonate produced by coral calcification is removed by erosion. There are three main mechanisms of reef erosion: physical erosion, chemical erosion (including rainwater dissolution), and bioerosion.
Physical erosion leads to the formation of grooves seen in most reefs from the action of waves moving rubble back and forth. This commonly develops into the ‘spur and groove’ structures common along reef fronts exposed to strong wave action.
Chemical erosion occurs primarily in shallow reefs exposed to rainwater, which is acidic. Rainwater dissolution of exposed reefs commonly results in ‘rill weathering’, a process that creates interlocking knife-like edges on the surface of limestone outcrops. Depending on the chemical composition of the limestone, some aerially exposed reefs endure this process much longer than others.
Bioerosion is a greatly under-recognised process, yet can be very active in shallow water where it not only erodes the limestone surface but prevents the growth of newly recruited corals and coralline algae on available substrates. Over thousands of years, the actions of many types of bioeroders such as urchins would probably be capable of keeping pace with slow sea-level falls, leaving no reef exposed above high tide level. There are many studies of the rate at which different organisms (such as sponges, urchins, limpets, chitons and parrot fish) ingest limestone (typically up to 18cm³ per animal per year for intertidal invertebrates such as sea urchins), however these cannot be reliably translated into rates at which these animals might plane-off whole reef surfaces.
Herbivory
The somewhat anthropomorphic notion that corals build reefs in order to have shallow substrates where they can out-compete macro-algae correctly implies that corals are the key structural components of reefs. However, herbivorous fish that eat these algae are the key functional components. If fish, and indeed other crucial algal grazers such as urchins, were removed from most reefs for just a few years, the corals would be smothered by algae. All these animal groups ultimately depend on the presence of healthy reef ecosystems.

Mixture of macroalgae and coral at the southern-most reef of the Indian Ocean. Houtman Abrolhos Islands, Western Australia. Photograph: Charlie Veron. Figure 19.
Zonation
Most accounts of coral reefs draw attention to the coral zones that they invariably create. These zones have been likened to zones of vegetation found on mountain slopes, as well they might, but on coral reefs they are mostly created by decreasing light and turbulence with increasing depth and, even in very clear water, this occurs within a depth range of less than 50 metres.

Zonation patterns. Oceanic reef showing a number of clearly delineated reef zones. From the right: deeper water zones leading to sharply outlined outer wave-swept crest, to outer reef flat, to inner sandy lagoon and beyond to the less exposed western edge. Middleton Reef, South-eastern Australia. Photograph: David Doubilet. Figure 20.
Contrary to popular belief, species diversity is not at its highest on the scenically attractive reefs found in clear oceanic water. It reaches a peak for any given region on fringing reefs protected from strong wave action, especially where the water is slightly turbid. This is partly because Acropora, the most speciose Indo-Pacific genus, seldom dominates these habitats. In wave-washed clear water habitats, a few species of Acropora may occupy most of the available space, whereas on more-sheltered turbid reefs, Acropora species are less spatially dominant, yet can occur with high local species diversity.
Reef flats and crests Calcareous algae thrive but corals are uncommon on parts of reefs exposed to extreme wave action. Small encrusting and submassive corals, mostly species of Isopora, Acropora and Pocillopora, may be present, typically developed in small crevices in the otherwise solid, wave-battered pavement of the reef crest. Where there is some protection and where the water is clear, Indo-Pacific reef crests and outer reef flats are often dominated by Acropora. In more sheltered areas coral diversity increases with massive and submassive corals in the genera Goniastrea, Favia, Favites, Platygyra and Porites and branching, encrusting and foliose Montipora spp. also common. Porites may form large ‘micro-atolls’, a characteristic feature of intertidal areas of many Indo-Pacific reef flats.
A somewhat analogous situation occurs in the Caribbean, although most genera are different and there are only two common species of Acropora.
On their inner margins, reef flats may be subject to very high water temperatures, and the build-up of terrigenous sediments and corals are increasingly replaced by macro-algae and seagrasses. Several coral species tend to specialize in these habitats, notably branching species of Montipora in the Indo-Pacific. In the Caribbean, this niche is filled by Manicina areolata and several other hardy species.
Shallow reef slopes Clear water reef slopes subject to moderate levels of wave action are often dominated by Acropora, notably several table-forming species in the Indo-Pacific and two elkhorn and staghorn species in the Caribbean. In more sheltered areas, and in more turbid conditions, massive and branching species of Porites and foliose Montipora and Turbinaria can dominate shallow Indo-Pacific reef slopes. In the Atlantic, these roles are filled by ecological equivalents in the genera Siderastrea, Diploria, Montastraea, Porites and Agaricia, among others.
Mid- to lower reef slopes and lagoons Below the major influence of wave energy, the structure of coral communities can become highly diverse, as this habitat is suitable for most species of coral. Alternatively, large sections of reef slope may be covered by a single species, as the relatively stable conditions are exploited by competitively-dominant corals. In the Indo-Pacific, columnar species of Goniopora or plating Pachyseris may fill this role, although many species that typically occur as small colonies can episodically form large monospecific stands or beds. Most coral species can survive on reef slopes to around 30m depth, after which species diversity diminishes with reducing illumination, until a few ‘mesophotic zone’ specialists (for further information see Mesophotic environments), notably in the genus Leptoseris, dominate. In the Indo-Pacific, a small number of specialized Acropora species also exploit this deep-water niche, although not in the Atlantic.
